

5. Пути и механизмы преобразования энергии в живых системах
5.1. Метаболизм. Катаболизм и анаболизм
Катаболизм – это процессы распада сложных веществ до более простых, сопровождающееся выделением энергии.
Анаболизм – процессы синтеза сложных молекул из более простых, сопровождающиеся поглощением энергии.
Совокупность всех катаболических и анаболических процессов, протекающих в организме – метаболизм.
5.2. Принципы биоэнергетики. АТФ – основной источник и аккумулятор энергии в организме. Макроэргические соединения. Фазы катаболизма основных питательных веществ в организме. Роль высокоэнергетических фосфатов в улавливании энергии. Субстратное и окислительное фосфорилирование
Принципы биоэнергетики. АТФ – основной источник и аккумулятор энергии в организме
Химическая термодинамика является теоретической основой биоэнергетики. Биоэнергетика занимается изучением энергетических превращений, сопровождающих биохимические реакции. Все эти превращения осуществляются в полном соответствии с первым и вторым началами термодинамики. Тем не менее, живой организм, как объект для термодинамических исследований, отличается целым рядом специфических особенностей в сравнении с системами, которые служат объектами изучения в технической и химической термодинамики. Из этих особенностей наибольшего внимания заслуживают следующие.
Живой организм представляет собой типично открытую систему, непрерывно обменивающуюся с окружающей средой веществом и энергией. В то же время для биологических систем применимо понятие стационарного состояния. В этом состоянии параметры в системе остаются постоянными, а скорость притока веществ и энергии равна скорости удаления их из системы. Такие системы не изменяют своих свойств во времени и сходны с системами, находящимися в равновесии.
Характер изменения энтропии, имеющий решающее значение при оценке процессов в неживых системах, в случае биологических систем имеет лишь подчиненное значение. Ведь все живые организмы высоко организованы и постоянно поддерживают свой уровень организации, поэтому для биологических систем справедливо неравенство Sокр. + Sсист. > 0 , а не Sсист > 0.
Все биохимические процессы, происходящие в клетках в условиях постоянства температуры и давления, при отсутствии значительных перепадов концентраций, резких изменений объема.
Небиологические системы могут совершать работу за счет тепловой энергии, биологические функционируют в изотермическом режиме и для осуществления процессов жизнедеятельности используют химическую энергию.
Стандартное состояние для биологических объектов отличается от стандартного состояния технических и химических систем. Так, стандартным состоянием химической системы является состояние, при котором концентрации всех веществ равны 1 моль/л (или для газов, входящих в систему, парциальные давления равны 1 атм). Для биологических систем это определение остается в силе за исключением концентрации ионов Н+, которая для стандартного состояния принимается равной 10-7 моль/л. Это соответствует нейтральной среде (рН 7), характерной для биологических объектов.
G0 - стандартная энергия Гиббса для химических систем;
G0 - стандартная энергия Гиббса для биологических систем.
Человек получает энергию за счет разложения органических веществ пищи. Органические вещества являются термодинамически нестабильными. Катаболические превращения этих веществ (распад или окисление) протекают с уменьшением свободной энергии. Такие процессы являются самопроизвольными (экзергоническими -G<0) и могут служить источником энергии для функционирования живой клетки. Все процессы, которые идут с увеличением свободной энергии (эндергонические -G>0), несамопроизвольные и должны быть сопряжены с экзергоническими процессами.
Жизненно важные процессы – реакции синтеза (т.е. анаболические процессы), мышечное сокращение, активный транспорт – являются эндергоническими процессами. Эндергонический процесс не может протекать изолированно. Такие процессы получают энергию путем химического сопряжения с реакциями окисления молекул пищевых веществ (катаболическими процессами), которые являются экзергоническими реакциями. Совокупность метаболических и анаболических процессов есть метаболизм.

Схематически такое сопряжение можно проиллюстрировать следующим образом. Пусть превращение метаболита (промежуточного соединения в цепи реакций) А в метаболит В сопровождается выделением свободной энергии. Оно сопряжено с другой реакцией - превращением метаболита С в метаболит D, которое может происходить лишь при поступлении свободной энергии.
Один из возможных механизмов состоит в образовании промежуточного соединения - общего для обеих реакций.
Примером являются дегидрогеназные реакции (реакции отщепления атомов водорода), промежуточным соединением, в которых является промежуточный переносчик атомов водорода.

Еще один пример сопряжения состоит в синтезе в ходе экзергонических реакций высокоэнергетического соединения, общего для многих процессов, и последующего включения этого соединения в эндергонические реакции, что обеспечивает передачу энергии. Таким общим для многих реакций соединением - энергетической валютой клетки - является аденозинтрифосфат (АТФ).
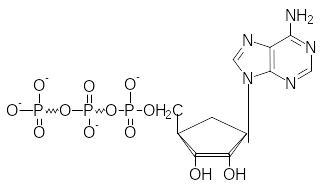
В молекуле АТФ две макроэргические связи, т.е. связи, при гидролизе которых высвобождается энергия. Это позволяет сопрягать большее число экзергонических реакций с большим числом эндергонических.
Главная роль АТФ в организме связана с обеспечением энергией многочисленных биохимических реакций. Являясь носителем двух высокоэнергетических связей, АТФ служит непосредственным источником энергии для множества энергозатратных биохимических и физиологических процессов. Всё это реакции синтеза сложных веществ в организме: осуществление активного переноса молекул через биологические мембраны, в том числе и для создания трансмембранного электрического потенциала; осуществления мышечного сокращения.
В организме АТФ является одним из самых часто обновляемых веществ, так у человека продолжительность жизни одной молекулы АТФ менее 1 мин. В течение суток одна молекула АТФ проходит в среднем 2000—3000 циклов ресинтеза (человеческий организм синтезирует около 40 кг АТФ в день), то есть запаса АТФ в организме практически не создаётся, и для нормальной жизнедеятельности необходимо постоянно синтезировать новые молекулы АТФ.