

Эволюционное учение
.pdfпозволяет выявлять направления возможной эволюции генома таких форм, т. е. выяснять их эволюционные взаимоотношения. На рис. 6.28 приведены хромосомные наборы ряда близких форм, которые могли возникнуть лишь в процессе последовательной полиплоидизации генома исходного типа.
Методологически важным результа- |
|
|
|
|
|
|
|
|
|
|
|||||||||||||
том проникновения генетики в эво- |
|
|
|
|
|
|
|
|
|
|
|||||||||||||
люционное |
учение |
стало |
широкое |
|
|
|
|
|
|
|
|
|
|
||||||||||
распространение |
|
экспериментальных |
|
|
|
|
|
|
|
|
|
|
|||||||||||
подходов |
|
в |
решении |
эволюционных |
|
|
|
|
|
|
|
|
|
|
|||||||||
задач, |
а |
также |
применение |
точного |
Рис. 6.28. |
Возникновение |
видов |
посредством |
|||||||||||||||
(сравнительно с тем, что есть в других |
|||||||||||||||||||||||
полиплоидизации. А—В—хромосомные наборы в |
|||||||||||||||||||||||
разделах |
|
биологии) |
аппарата |
гене- |
|||||||||||||||||||
|
первой метафазе мейоза трех видов пшениц |
||||||||||||||||||||||
тических |
понятий. |
|
|
|
|
|
|
|
Triticum |
aegilopoides, |
п=1\ |
Т. |
aestivum, /г=21; |
||||||||||
Разработка |
|
современного |
|
эволю- |
Т. dicoccoides, п= 14; Г — Е — то же, для |
видов |
|||||||||||||||||
ционного учения немыслима вне гене- |
хризантем: Chrysanthemum makinoi, п—9; |
Ch. |
|||||||||||||||||||||
indicum, |
р= 18; |
Ch. |
japonense, |
n= 27 |
(no |
||||||||||||||||||
тических подходов и методов. Раз- |
|
|
|
А. Мюнтцингу, |
1967) |
|
|
||||||||||||||||
витие |
популяционной |
морфологии, |
|
|
|
|
|
|
|
|
|
|
|||||||||||
фено- и геногеографии, микросисте- |
нуклеотидов (в ДНК и РНК) и |
||||||||||||||||||||||
матики, |
кариосистематики |
и |
|
других |
аминокислот (в белках). На современ- |
||||||||||||||||||
«промежуточных» |
дисциплин |
(опреде- |
ном этапе развития молекулярной био- |
||||||||||||||||||||
ляющих в известной мере современ- |
логии можно анализировать число раз- |
||||||||||||||||||||||
ный |
этап |
развития |
эволюционного |
личий в последовательностях элемен- |
|||||||||||||||||||
учения) |
наглядно |
|
иллюстрирует |
эту |
тов нуклеиновой кислоты или белка |
||||||||||||||||||
ведущую |
роль |
генетики. |
|
|
|
|
|
разных видов, судить по этому по- |
|||||||||||||||
Методы |
биохимии и |
молекулярной |
казателю о степени их отличий* По- |
||||||||||||||||||||
биологии. Уже Ч. Дарвин сделал |
скольку |
каждая замена |
аминокислоты |
||||||||||||||||||||
успешные |
попытки |
использовать |
в белке может быть связана с |
||||||||||||||||||||
биохимические показатели для уста- |
изменением одного, двух или трех |
||||||||||||||||||||||
новления |
систематической |
принадлеж- |
нуклеотидов |
в |
молекуле |
ДНК, |
на |
||||||||||||||||
ности тех или иных форм. Однако |
ЭВМ |
можно |
вычислить |
максимальное |
|||||||||||||||||||
только начиная с середины XX в. |
и |
минимальное |
число |
нуклеотидных |
|||||||||||||||||||
биохимические и, в частности, моле- |
замен, |
необходимых |
для |
замещения |
|||||||||||||||||||
кулярнобиологические |
методы |
|
вышли, |
аминокислот |
в белке. |
|
|
|
|
||||||||||||||
пожалуй, на передовые рубежи в изу- |
|
Получаемая таким образом инфор- |
|||||||||||||||||||||
чении эволюционного процесса. По ана- |
мация поддается дальнейшей коли- |
||||||||||||||||||||||
логии |
с |
изучением |
эволюции |
органов |
чественной |
оценке: |
при |
сравнении |
|||||||||||||||
и онтогенеза |
в |
XIX — начале |
XX |
в. |
ряда |
организмов |
можно |
установить |
|||||||||||||||
возникло |
целое |
направление |
изучения |
и степень различий между ними по |
|||||||||||||||||||
«молекулярной |
эволюции». |
Здесь |
мы |
аминокислотам или нуклеотидам. Дру- |
|||||||||||||||||||
рассмотрим лишь некоторые из мето- |
гое |
важное |
преимущество |
изучения |
|||||||||||||||||||
дов молекулярной |
|
биологии, |
широко |
эволюции методами молекулярной био- |
|||||||||||||||||||
применяющихся |
ныне |
для |
изучения |
логии — возможность сравнения сколь |
|||||||||||||||||||
эволюционного |
процесса. |
|
|
|
|
угодно далеких организмов — растений |
|||||||||||||||||
В ы я с н е н и е |
|
с т р о е н и я |
н у - |
и животных, грибов и микроорга- |
|||||||||||||||||||
к л е и н о в ы х |
к и с л о т |
и б е л к о в . |
низмов, На рис. 6.29 показан результат |
||||||||||||||||||||
На молекулярном уровне процесс эво- |
изучения филогенетических отношений |
||||||||||||||||||||||
люции |
связан |
с |
изменением |
состава |
20 |
разных организмов |
на |
основе |
оп- |
||||||||||||||
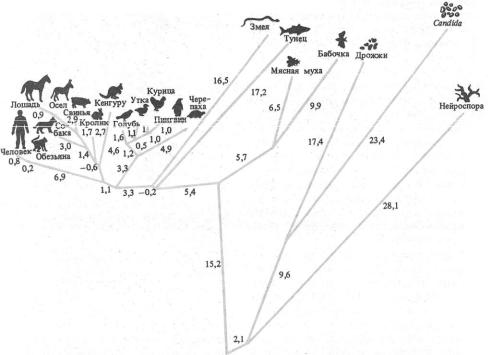
92 Часть 2. Органическая эволюция как объективный процесс
Рис. 6.29. Филогенетические отношения между разными организмами, построенные на основании изменений белка цитохрома-С.
Цифры—число мутаций (по Ф. Айяла, 1981)
ределения минимального числа воз- |
уровне является оцен!& эволюцион- |
||||||||||||
можных нуклеотидных различий меж- |
ных изменений по степени сходства |
||||||||||||
ду генами, |
кодирующими**' синтез |
первичной структуры нуклеиновых кис- |
|||||||||||
белка цитохрома-С. Можно видеть, что |
лот у различных групп организмов |
||||||||||||
в общем эти данные хорошо совпа- |
посредством |
гибридизации |
ДНК. Хотя |
||||||||||
дают с выводами классической систе- |
около 90% ДНК не кодирует белков, |
||||||||||||
матики. Конечно, данным, полученным |
тем не менее изучение ДНК позволяет |
||||||||||||
на основании изучения одного лишь |
оценивать филогению генов. Нуклео- |
||||||||||||
белка, нельзя |
придавать абсолютного |
тидные |
последовательности |
позволяют |
|||||||||
значения: более точно филогения будет |
судить |
об |
эволюции |
генов |
точнее, |
||||||||
выяснена |
при |
сравнении |
аналогичных |
чем другие методы молекулярной био- |
|||||||||
схем, построенных по многим раз- |
логии. Расщепленная на отдельные ни- |
||||||||||||
личным белкам. Это и понятно: не |
ти ДНК одного организма «гибриди- |
||||||||||||
может быть ни одного универсального |
зируется» с молекулами ДНК другого |
||||||||||||
белка, имеющего |
одинаковое |
значение |
вида и в зависимости от того, |
||||||||||
(а следовательно, и менявшегося с |
насколько |
различаются |
последователь- |
||||||||||
одинаковой скоростью) |
в |
эволюции |
ности нуклеотидов, гибридизация за- |
||||||||||
всех групп организмов. Какие-то белки |
хватывает |
|
большие |
или |
|
меньшие |
|||||||
должны |
быть |
|
более |
стабильными, |
участки нитей ДНК. Этим достигает- |
||||||||
какие-то — менее |
в |
каждой |
крупной |
ся количественная оценка эволюцион- |
|||||||||
группе. |
|
|
|
|
|
|
ных изменений, происшедших со срав- |
||||||
Другим методом изучения эволю- |
ниваемыми видами. ДНК человека ока- |
||||||||||||
ционного |
процесса |
на |
молекулярном |
зывается |
гомологичной |
ДНК |
макаки |
||||||
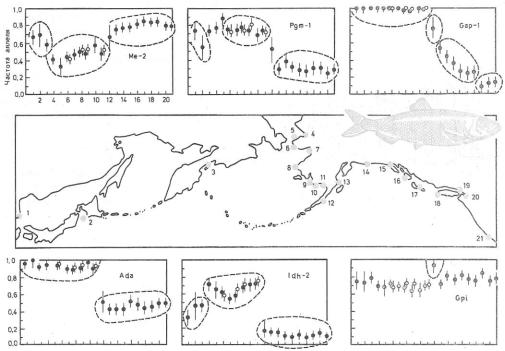
Рис. 6.30. Пример феногеографического анализа по частотам аллозимов (Ме-2, Pgm-1, Gap-1,
Ada, |
ldh-2, |
Gpi) популяционной |
структуры тихоокеанской |
сельди Clupea |
pallasi |
(по В. Грант, |
|||||||||||
|
|
|
|
|
|
|
|
|
|
Ф. Уттер, |
1984): |
|
|
|
|
||
|
|
|
|
|
|
|
|
|
|
1—21—места сбора |
|
|
|
|
|||
на 66%, |
быка — на |
28, |
крысы — на |
ДНК. Скорость эволюции макромоле- |
|||||||||||||
17, лосося — на 8, бактерии кишечной |
кул в разных филогенетических ли- |
||||||||||||||||
палочки — на |
2%. |
|
|
|
|
|
|
ниях растений оказывается далеко не |
|||||||||
Изучение |
особенностей |
эволюции |
одинакова. Так, темпы накопления из- |
||||||||||||||
на молекулярном уровне привело к идее |
менений в нуклеотидных последова- |
||||||||||||||||
наличия молекулярных часов, не только |
тельностях у линий, ведущих к цветко- |
||||||||||||||||
отражающих, но и регулирующих эво- |
вым, более чем в 10 раз выше, чем в |
||||||||||||||||
люционный |
процесс. |
Такие |
расчеты |
линии высших нецветковых. При изу- |
|||||||||||||
основаны на предположениях о на- |
чении генов актинов было выявлено, |
||||||||||||||||
коплении изменений в информацион- |
что различия между близкими видами |
||||||||||||||||
ных |
макромолекулах |
с |
постоянной |
у растений превышают таковые разных |
|||||||||||||
скоростью. Однако оказалось, что для |
классов у животных. Поэтому ско- |
||||||||||||||||
растений, отличающихся в целом ис- |
рость хода молекулярных часов ока- |
||||||||||||||||
ключительно высокой скоростью эво- |
зывается резко различной в разных |
||||||||||||||||
люции |
первичной |
|
структуры |
ДНК, |
группах, а также, по-видимому, в раз- |
||||||||||||
концепция молекулярных часов не- |
ные |
геологические |
|
эпохи |
(см. также |
||||||||||||
приемлема: ДНК |
растений |
из |
разных |
гл. 15, 20). |
|
|
|
||||||||||
семейств |
отличается |
в |
такой |
же сте- |
|
|
|
||||||||||
Иммунологические методы. Биохи- |
|||||||||||||||||
пени, |
как |
ДНК |
животных |
разных |
|||||||||||||
мия |
располагает специальными мето- |
||||||||||||||||
классов. У |
растений |
в |
ходе |
эволюции |
|||||||||||||
дами |
исследования, |
позволяющими с |
|||||||||||||||
очень быстро |
могут меняться и повто- |
||||||||||||||||
большой |
степенью |
|
точности выяснить |
||||||||||||||
ряющиеся |
|
и |
уникальные |
|
последова- |
|
|||||||||||
|
|
«кровное |
родство» |
разных групп. При |
|||||||||||||
тельности |
|
нуклеотидов |
в |
|
молекулах |
||||||||||||
|
|
сравнении белков |
крови |
используется |
|||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|||||||
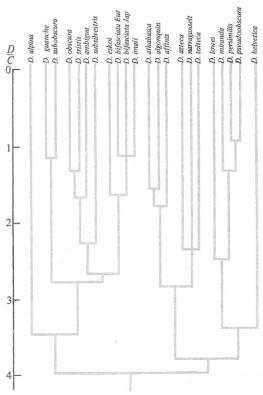
94 |
Часть 2. Органическая эволюция как объективный процесс |
|
(птицы оказываются филогенетически ближе к крокодилам, чем крокодилы к змеям). Показаны глубокие серологические различия между хвостатыми и бесхвостыми амфибиями; найдены черты сходства по реакции преципитации между хордовыми и иглокожими и т. д. Подобные методы исследований применимы и к растениям.
Биохимический полиморфизм. Начиная с 60-х годов в практику микроэволюционных исследований широко входит метод изучения вариаций белков, обнаруживаемых с помощью электрофореза (Р. Левонтин). С помощью этого метода можно с большей точностью, чем прежде, определять уровень генетической изменчивости в популяциях, а также степень сходства и различия между популяциями по сумме таких вариаций.
Различия между популяциями по биохимическим показателям можно наглядно определять на карте (рис. 6.30) либо на дендрограмме, построенной по коэффициентам сходства и различий (рис. 6.31).
|
|
|
|
|
|
|
|
|
|
|
По |
частотам |
электроморф |
можно |
||||||||
Рис. 6.31. Пример построения схемы филогене- |
исследовать |
и |
внутриг^опуляционную |
|||||||||||||||||||
тических отношений в виде дендрограммы на |
пространственно-генетическую струк- |
|||||||||||||||||||||
основании данных электрофореза белков: взаимо- |
туру и выявлять динамику этой струк- |
|||||||||||||||||||||
отношения |
между _ видами |
дрозофил |
группы |
туры, |
т. е. исследовать |
|
самые |
началь- |
||||||||||||||
D. pseudoobscura. Д/С—среднее число амино- |
ные |
стадии |
процесса |
микроэволюции |
||||||||||||||||||
кислот, по которым различаются ферменты раз- |
||||||||||||||||||||||
ных |
видов |
(из |
С. М. Гершензона, |
1979) |
|
(см. гл. 8, 9). |
|
|
|
|
|
|
||||||||||
|
|
|
|
|
|
|
|
|
|
|
При |
биохимических |
сравнениях |
|||||||||
способность |
организмов |
|
вырабатывать |
ныне |
|
живущих |
организмов |
показано, |
||||||||||||||
(в |
ответ |
на |
введение |
в |
кровь |
что |
ранние |
метаболические |
звенья |
|||||||||||||
чужих |
|
белков) |
антитела, |
которые |
синтеза стеринов, каротиноидов, вто- |
|||||||||||||||||
затем можно выделить из сыворотки |
ричных метаболитов, реакции глико- |
|||||||||||||||||||||
крови и определить, при каком раз- |
лиза |
и |
дыхания |
у |
многих растений |
|||||||||||||||||
ведении эта сыворотка будет реаги- |
и |
животных |
сходны и |
|
не |
нуждаются |
||||||||||||||||
ровать с другой, сравниваемой сыво- |
в |
участии |
кислорода. |
|
Завершающие |
|||||||||||||||||
роткой |
(реакции |
преципитации). Такой |
же стадии многих метаболических пу- |
|||||||||||||||||||
анализ показал, например, что бли- |
тей, особенно у высокоорганизованных |
|||||||||||||||||||||
жайшие родственники человека — выс- |
существ, |
протекают |
с |
|
его |
участием. |
||||||||||||||||
шие |
человекообразные |
|
обезьяны, а |
Это свидетельствует об их эволюци- |
||||||||||||||||||
наиболее |
далекие среди |
приматов — |
онной молодости |
и |
сложении в аэроб- |
|||||||||||||||||
ной |
фазе истории |
Земли. |
Таким об- |
|||||||||||||||||||
лемуры. |
|
Обнаружено |
серологическое |
|||||||||||||||||||
|
разом, |
|
с |
помощью |
биохимических |
|||||||||||||||||
сходство |
крови |
хищных |
млекопитаю- |
|
||||||||||||||||||
методов |
(и |
в том числе |
молекулярно- |
|||||||||||||||||||
щих |
с |
ластоногими, обнаружена |
бли- |
|||||||||||||||||||
биологических) |
можно |
исследовать |
||||||||||||||||||||
зость птиц |
к крокодилам |
и |
черепахам |
|||||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|||||||||||
эволюционные процессы самого разного масштаба. Несомненно, что в ближайшие годы арсенал этих методов должен существенно расшириться, учитывая значительное развитие исследований в этой области.
Методы моделирования |
эволюции. |
В последние десятилетия |
появилась |
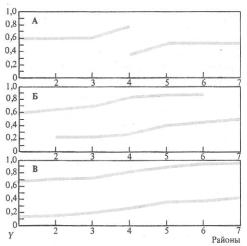
возможность моделировать эволюционный процесс'. Конечно, сложность организации жизни такова, что пока невозможно надежное моделирование развития экосистемы или целой группы, но влияние отдельных эволюционных факторов на популяцию и вид в целом поддаются моделированию. Пока такое моделирование неизбежно связано с принятием целого ряда упрощающих условий (например, рассматриваются обычно лишь двулокусные генетические системы, принимается равновероятной возможность скрещивания особей в популяции друг с другом и т. п. упрощения, далекие от природных ситуаций). Но и при таких ограничениях моделирование иногда помогает уже сейчас решать ряд эволюционных задач. Моделирование эволюционного процесса позволяет, в частности, исследовать возможные влияния отдельных факторов на ход эволюции, их взаимодействие, выяснить в некоторых случаях возможные направления и результаты процесса эволюции (рис. 6.32).
При применении быстродействующих ЭВМ с большой памятью возникает возможность как бы резко ускорить «процесс эволюции» и составить прогноз развития эволюционных событий при различном сочетании условий.
Особое внимание при моделировании процесса эволюции приходится уделять выбору необходимых и доста-
1 Как известно, под моделированием понимается метод опосредованного оперирования объектом, при котором исследуется не сам объект, а вспомогательная система (модель), способная заменять объект на определенных этапах познания; такое исследование в конечном счете может дать информацию о самом моделируемом объекте.
Рис. 6.32. Пример моделирования эволюции: распространение признака Y (форма рыла: острорылость—тупорылость) в популяциях ленка (Brachymystax) в Азии: А—стадия расхождения признаков в районе первого контакта; Б— промежуточное состояние; В—возникшее стационарное состояние по 'данной паре признаков
(из М. В. Мины, 1985)
точных исходных предпосылок. Можно заложить в машину и «проиграть»
различнейшие |
варианты |
взаимодейст- |
|||
вия |
тех |
или |
иных |
эволюционных |
|
сил |
в той |
или |
иной |
«обстановке», |
|
но |
если |
упустить какой-либо один |
|||
из ведущих компонентов, то результаты моделирования могут получиться обратные реально существующим в природе. Определить набор необходимых и достаточных предпосылок для создания модели — дело очень сложное. Такие работы целесообразно организовывать только в тесном контакте математиков и биологов.
Другие методы изучения эволюции. На основании данных полученных сравнительной физиологией, можно проследить эволюционное развитие тех или иных функций. Так, по обмену веществ грибы ближе стоят к животным, чем к растениям (А. Н. Белозерский), что служит еще одним основанием для выделения их в самостоятельное царство.
Значительный интерес для выяснения тонких особенностей протекания
96 |
|
Часть 2. Органическая эволюция как объективный |
процесс |
|
|||
процесса эволюции представляет изуче- |
му отбору и пр.), а также расшиф- |
||||||
ние поведения |
животных |
(этология), |
ровка так называемых «природных экс- |
||||
на что много внимания обращал Ч. Дар- |
периментов», постоянно |
возникающих |
|||||
вин. У близких видов, недавно дивер- |
в природе4 (возникновение и нару- |
||||||
гировавших от общего предка, сохра- |
шение разного |
рода |
эволюционных |
||||
няется |
(или |
рекапитулирует |
в про- |
барьеров между популяциями, вселе- |
|||
цессе |
онтогенеза) множество |
общих |
ние новых форм и т. д.). |
||||
мелких |
черт |
поведения |
(К. |
Лоренц, |
Много микроэволюционных ситуа- |
||
Н.Тинберген и др.). Некоторые ций можно исследовать и в наши
характерные древние черты поведения у птиц и млекопитающих могут быть не менее важными показателями происхождения от общих предков, чем гомологичные органы или другие классические сравнительно-анатомические доказательства.
В некоторых случаях эффективным оказывается использование паразит ологического метода изучения эволюции. Многочисленными исследованиями доказано, что эволюция паразитов и хозяев протекает сопряженно. Известно также, что в некоторых группах паразиты оказываются специфическими для видов, родов или семейств. Поэтому по присутствию определенных паразитов порой можно с большой точностью судить о филогенетических связях видов-хозяев этих паразитов.
По-видимому, любая биологическая дисциплина обладает своим специфическим набором методов изучения эволюционного процесса и достаточно полное их перечисление практически невозможно, да и не нужно в рамках курса дарвинизма.
6.3. Особенности изучения микро-
и макроэволюции
Для анализа микроэволюции —
процессов, текущих внутри вида на протяжении сравнительно коротких отрезков времени,—применяют комплекс попу ляционных методов. Для изучения микроэволюции характерно также проведение экспериментов (не только генетических, например гибридологического анализа, но и экологических, таких, как эксперименты по борьбе за существование, естественно-
дни при анализе результатов антропогенного влияния на живую природу. При этом прежде единые ареалы популяций дробятся на части, возникают новые или исчезают старые изоляционные барьеры, изменяется (порой резко) давление эволюционных факторов. Интересной задачей оказывается достаточно точный учет таких изменений и реакций на них природных популяций тех или иных видов. К сожалению, в большинстве случаев мы не успеваем зафиксировать исходные состояния популяций до начала антропогенного воздействия, и лишаемся тем самым возможности оценить величину и скорость происшедших впоследствии микроэволюционных изменений.
Однако уже сейчас накопилось не-
мало данных |
по микроэволюционным |
|
изменениям, |
например, |
происшедшим |
со многими |
видами и |
популяциями |
в результате случайной или направленной акклиматизации растений, беспозвоночных, рыб, птиц и млекопитающих. Есть много данных по возникновению устойчивости членистоногих и позвоночных к используемым против них пестицидам, а растений — к гербицидам. Есть интересные данные повышения толерантности растений к содержанию в среде тяжелых металлов и других загрязняющих веществ. Все это заслуживает тщательного анализа с позиций исследования процессов микроэволюции. Популяционные методы исследования сейчас проникают в палеонтологию, биохимию, этологию и многие другие науки.
Для анализа макроэволюции — процессов, текущих на протяжении десятков и сотен миллионов лет,—
Глава 6. Методы .изучения эволюции 97
применяются не менее разнообразные методы. Среди них на одном из первых мест стоит комплекс палеонтологических методов: сбор материала и восстановление облика ископаемых форм, их экологии, биогеографии, анализ веществ биогенного происхождения, изучение микроскопических ископаемых организмов и т. д. Для анализа макроэволюции успешно используется изучение современных форм. Сравни- тельно-морфологические методы изучения эволюции служат для установления родства форм по степени сходства в их строении. Эмбриологический
метод изучения эволюции основан на сравнении ранних этапов онтогенеза разных организмов; при этом иногда удается выявить предковые признаки, исчезающие на более поздних стадиях развития1. Анализ белков и нуклеиновых кислот позволяет устанавливать сходство и различие разных организмов; эти и другие биохимические методы чаще всего применяются при молекулярно-биологическом подходе к изучению эволюционного процесса. Биогеографические методы позволяют восстанавливать историю распространения, расцвета и увядания разных групп в пространстве биосферы.
Паразитологические методы, основанные на теснейшем эволюционном единстве паразитов со своими хозяевами, позволяют в ряде случаев по паразитофауне строить обоснованные предположения об эволюции отдельных видов, родов, семейств хозяев. Изучение тонких особенностей поведения животных — этологические методы — позволяют устанавливать родство между группами на основании гомологии генетически обусловленных элементов поведения. Физиологические методы изучения эволюции, основанные на сопоставлении особенностей обмена веществ и проявления функциональных
1 Совокупность сравнительно-анатомических, сравнительно-эмбриологических и палеонтологических методов в изучении эволюции со времен Э. Геккеля называют методом тройного параллелизма.
4 Зак. 2179
особенностей у разных организмов, также помогают воссоздавать пути происхождения тех или иных форм.
6.4. Необходимость комплексного подхода к изучению эволюции
Из кратко описанных в этой главе основных методов изучения эволюционного процесса вытекает один принципиально важный вывод: нет ни одного «абсолютного» или совершенного метода для изучения особенностей протекания процесса эволюции.
Наши представления об эволюции, опиравшиеся длительное время на классические методы, существенно расширяются с помощью новейших методов молекулярной биологии. Изучение процесса видообразования, сначала проводившееся только на основе морфологических и биогеографических методов, существенно дополнено генетическими, физиологическими, а в последнее время и биохимическими подходами. Изучение процессов эволюции экосистем, проводившееся на основе восстановления вымерших флор и фаун, а также палеоэкологии, существенно расширяется в связи с использованием методов математического моделирования, термодинамическими и информационными подходами. Число таких примеров можно увеличить.
Показательна ситуация, сложившаяся в 80-е годы XX в. при изучении особенностей процесса микроэволюции с применением морфофизиологических и биохимических методов. В ряде случаев результаты таких исследований, например, при определении сходства
иразличия популяций полностью
совпадали, в других — не совпадали и даже давали прямо противоположные оценки. В конце концов после длительных споров о преимуществах того или другого подходов удалось показать, что эти данные и не стоит сравнивать — они отражают существенно разные стороны протекания процесса эволюции, и получаемые таким об-
98 |
|
|
Часть |
2. |
Органическая |
|
эвОЛЮЦИЯ |
как |
объективный |
процесс |
|
|
|||||||||||
разом характеристики являются до- |
предстает не в виде правдоподобной |
||||||||||||||||||||||
полняющими |
одна |
другую |
|
(в |
соответ- |
гипотезы, а в качестве неопровержи- |
|||||||||||||||||
ствии |
с |
известным |
общенаучным |
мого факта, |
подтверждаемого |
суммой |
|||||||||||||||||
принципом дополнительности Н. Бора). |
наших знаний из разных областей |
||||||||||||||||||||||
Несмотря на сравнительно широкий |
естественных наук. |
|
|
|
|||||||||||||||||||
спектр уже известных методов, было бы |
|
|
В комплексе разные методы позво- |
||||||||||||||||||||
неверным |
считать, что |
возможности |
ляют |
изучать |
эволюционный |
процесс |
|||||||||||||||||
изучения эволюции в этом направле- |
от происхождения крупных групп и |
||||||||||||||||||||||
нии исчерпаны. Параллельно с про- |
экосистем |
|
до |
возникновения |
новых |
||||||||||||||||||
грессом |
|
в |
конкретных |
|
дисциплинах |
популяций |
и |
видов. |
|
|
|
||||||||||||
формируются все новые и новые мето- |
|
|
Сопоставление различных подходов |
||||||||||||||||||||
ды изучения эволюции. Только за |
к изучению эволюции в XIX и XX вв. |
||||||||||||||||||||||
последние 15—20 лет возникли такие |
показывает, что постоянно |
происходит |
|||||||||||||||||||||
ныне широко распространенные мето- |
углубление всех классических |
методов |
|||||||||||||||||||||
ды исследования |
|
процесса |
эволюции, |
исследования и развитие новых. Важ- |
|||||||||||||||||||
как изучение биохимического полимор- |
ная |
черта |
использования |
различных |
|||||||||||||||||||
физма, |
|
популяционная |
|
морфология, |
методов изучения эволюции — их вза- |
||||||||||||||||||
цитогенетические |
|
и |
кариологические |
имодополняемость: |
методы |
палеонто- |
|||||||||||||||||
методы, методы молекулярной биоло- |
логии органически сочетаются не толь- |
||||||||||||||||||||||
гии. Факты свидетельствуют, что сей- |
ко с морфологическими или биогеогра- |
||||||||||||||||||||||
час идет по существу активнейшее |
фическими, |
но |
и |
с |
биохимическими |
||||||||||||||||||
обновление и расширение фронта эво- |
и |
|
генетическими; эмбриологические |
||||||||||||||||||||
люционных |
исследований, |
последст- |
подходы |
естественно |
сочетаются не |
||||||||||||||||||
вием чего должно быть более глубо- |
только с морфологическими, но с |
||||||||||||||||||||||
кое понимание закономерностей про- |
физиологическими |
и |
биохимическими |
||||||||||||||||||||
цесса эволюции. |
|
|
|
|
|
|
|
|
|
и |
т. д. |
|
|
|
|
|
|
|
|
||||
Эволюция |
— |
историческое |
развитие |
|
|
|
|
|
|
|
|
|
|
|
|||||||||
живого |
на Земле |
— в настоящее время |
|
|
|
|
|
|
|
|
|
|
|
||||||||||
|
|
|
Наиболее общее и специфическое свойство |
|
живого — |
|
|||||||||||||||||
|
|
способность |
|
к |
|
самовоспроизведению |
на |
основе |
матрич- |
|
|||||||||||||
|
|
ного принципа. Эта способность вместе с другими осо- |
|
||||||||||||||||||||
|
|
бенностями |
|
живых |
существ |
определяет |
существование |
|
|||||||||||||||
|
|
четырех |
основных |
уровней |
организации |
живого |
(молеку- |
|
|||||||||||||||
|
|
лярно-генетический, |
онтогенетический, |
|
популяционно-ви- |
|
|||||||||||||||||
|
|
довой |
|
и |
биогеоценотический). |
|
|
|
|
|
|
|
|
|
|
||||||||
|
|
|
Все уровни организации жизни находятся в сложном |
|
|||||||||||||||||||
|
|
взаимодействии |
как |
части |
единого |
целого. В то |
же |
время, |
|
||||||||||||||
|
|
на каждом уровне действуют свои закономерности, |
|
||||||||||||||||||||
|
|
определяющие |
особенности |
эволюции всех форм органи- |
|
||||||||||||||||||
|
|
зации живого. Специфические свойства жизни |
обеспечи- |
|
|||||||||||||||||||
|
|
вают не только воспроизведение себе подобных |
|
(наследст- |
|
||||||||||||||||||
|
|
венность), но и необходимое для эволюции изменение |
|
||||||||||||||||||||
|
|
самовоспроизводящихся |
структур |
|
(изменчивость). |
|
|
|
|||||||||||||||
|
|
|
Огромное число фактов из разных областей биологии |
|
|||||||||||||||||||
|
|
свидетельствует, что процесс эволюции реально осуществ- |
|
||||||||||||||||||||
|
|
лялся |
|
на |
|
протяжении |
|
нескольких |
миллиардов |
лет |
|
||||||||||||
|
|
существования Земли. Разнообразные методы изучения |
|
||||||||||||||||||||
|
|
эволюционного |
|
|
процесса |
|
позволяют |
|
выяснить |
сущест- |
|
||||||||||||
|
|
венные |
|
черты |
эволюции |
|
любой |
группы |
живых |
|
организмов |
|
|||||||||||
|
|
|
Г л ав а 6. |
Методы .изучения эволюции |
|
99 |
|||||
в |
прошлом, |
восстанавливать |
|
пути |
в направлении |
развития |
|||||
их |
|
целых |
сообществ, |
воссоздавать |
общую |
карти- |
|||||
ну |
эволюционного |
развития |
больших |
пространств |
на- |
||||||
шей |
планеты. |
Палеонтологические, |
биогеографические, |
||||||||
морфологические, |
эмбриологические |
и |
другие |
методы |
|||||||
исследования, используемые в совокупности, дают возмож- |
|||||||||||
ность |
проследить |
|
развитие |
жизни |
как планетарного |
||||||
явления. |
Данные |
|
молекулярной |
биологии, |
биохимии, |
||||||
биогеоценологии позволяют создать модель первых этапов |
|||||||||||
возникновения жизни |
и |
образования |
биосферы |
Земли. |
|||||||
Существует немало методов для изучения протекания эволюционного процесса и в меньших масштабах — микроэволюции.
Эволюция жизни на Земле — неоспоримый факт, В чем же причины эволюции живого, каковы движущие силы и закономерности этой эволюции? Рассмотрение и анализ
этих |
вопросов составляет основное содержание современ- |
ной |
теории эволюции. |
1. Сопоставьте предполагаемые пути возникновения жиз-
2» Почему приходится говорить о неизбежности эво-
3. Перечислите характерные особенности жизни, выделяя наиболее важные для протекания процесса эволюции.
5. Охарактеризуйте основные уровни организации жизни на Земле и докажите их несводимость друг к другу.
«Солярис»? |
^ |
7. Как можно |
доказать объективность процесса эволю- |
8. В чем состоит существо метода тройного параллелизма и значение этого и подобных методологических подходов для решения спорных .вопросов филогенеза отдельных групп (с примерами)?
чения макро- и микроэволюционных процессов?
ности Н. Бора к изучениюУэволюции?
бабочка, сосна. Какими методами наиболее надежно
(родство) этих форм?
4 s
100Часть 2. Органическая эволюция как объективный процесс
13.Можно ли с помощью методов молекулярной биологии изучать эволюционные процессы внутривидового масштаба?
14.Можно ли палеонтологическими методами изучать микроэволюцию? Какова разрешающая сила современных палеонтологических методов?
15.Какими биогеографическими методами можно изучать процессы микроэволюции?
16.Как доказать, что сравниваемые структуры гомо-
логичные или аналогичные в случае их значительного внешнего сходства (например, вибриссы у млекопитающих и щетинки на теле насекомых?, колючки у шиповника и колючки у барбариса?).
17. В чем принципиальное отличие рудиментов от атавизмов?
18. Какие немногие особенности организации живого на Земле необходимы и достаточны для моделирования процесса эволюции?
19.Можно ли предсказать направления эволюции жизни на Земле в будущем, опираясь на ее результаты в прошлом? Каковы здесь предполагаемые подходы и перспективы?
20.Перечислите характерные черты эволюции растений и
животных, выделяя общие и специфические моменты.
21.Покажите взаимосвязь эволюции растений и животных на примере развития жизни в каком-либо геологическом периоде.
22.Оцените вклад методов разных наук в изучение процесса эволюции и возможные изменения в этом направлении в будущем.
23.Какова связь развития методов изучения эволюции с доказательствами объективности эволюционного процесса в живой природе.