

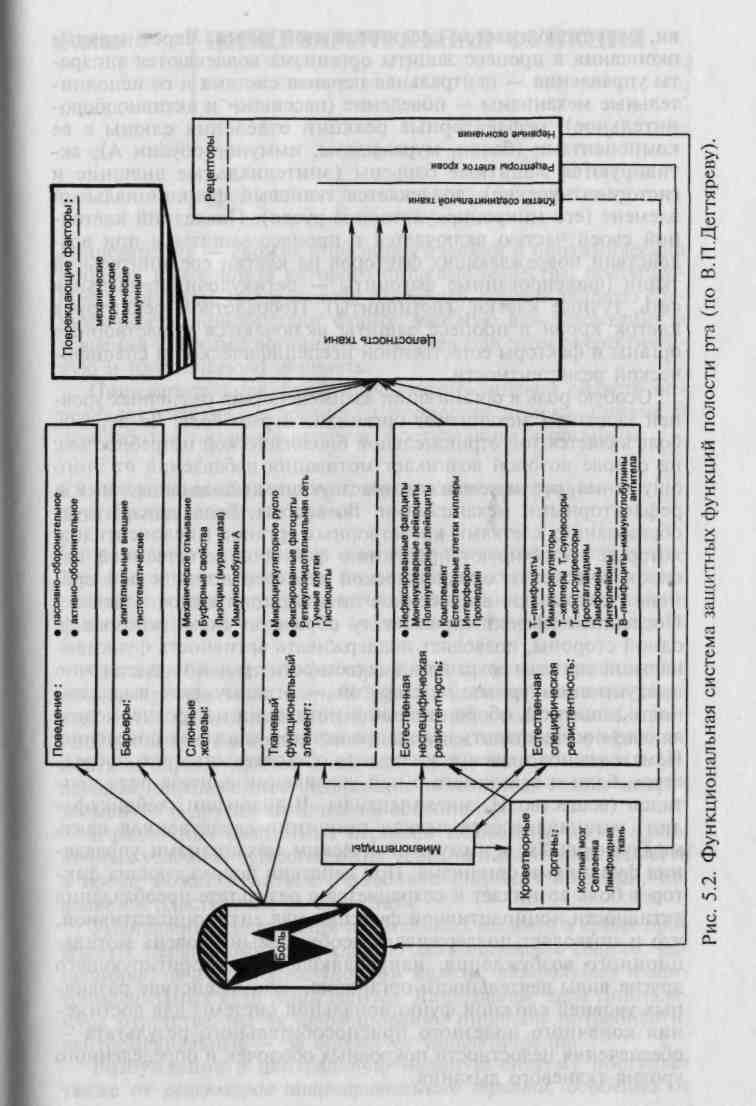
5.2.5. Факторы специфической резистентности
Иммунитет — способ защиты организма от живых тел и веществ, несущих на себе признаки генетической чужеродности. Иммунитет обеспечивается совместным функционированием
двух популяций лимфоцитов: тимусзависимых (Т-лимфоци-ты) и не зависящих в своем развитии от вилочковой железы (В-лимфоциты). Взаимодействие Т- и В-лимфоцитов и совместная их работа с макрофагами обеспечивают всю гамму иммунологических реакций, развивающихся в ответ на воздействие генетически чужеродных субстанций экзогенного и эндогенного происхождения. Этому способствует и появление ощущения боли при повреждении тканей, а также в процессе развития воспалительных реакций. Современными исследованиями показано, что переживание чувства боли способствует выработке веществ пептидергической природы — миелопепти-дов, стимулирующих рост и созревание лимфоцитов.
Лимфоциты обеспечивают клеточную и гуморальную формы иммунитета. В результате дифференцировки клеток лимфоциты приобретают распознающие структуры — рецепторы антигенов, биологически активных веществ — гистамина, про-стагландина Е, эндорфинов, лимфокинов. Биологически активные вещества оказывают опосредованное влияние на иммунные реакции через эти рецепторы.
Клеточный иммунитет включает распознавание антигена Т-лимфоцитами, активацию последних, трансформацию, усиление функциональной активности, в том числе лимфокин-продуцирующей и цитотоксической. Доминирующая роль в активации, пролиферации и дифференцировке Т-лимфоцитов принадлежит медиаторам, особенно интерлейкинам, в частности интерлейкину-1. Этот гормоноподобный монокин, продуцируемый активированными макрофагами, осуществляет дополнительный сигнал, необходимый для активации Т-лимфоцитов. В результате воздействия антигенных и дополнительных сигналов на лимфоциты происходит их дифференцировка с селекцией клонов лимфоцитов, специфичных для определенного антигена, а также пролиферация эффекторных клеток.
Т-лимфоциты делят на Т-эффекторы и иммунорегулятор-ные Т-лимфоциты.
Т-эффекторы (киллеры) характеризуются цитотоксической активностью, определяемой клетками, антителами, комплементом. Антигенспецифические Т-лимфоциты после предварительного контакта с антигеном приобретают специфичность к нему — становятся сенсибилизированными. Они повреждают клетки-мишени с помощью собственного рецептор-ного аппарата в несколько этапов: короткой фазы узнавания, летального удара и дезинтеграции.
Иммунорегуляторные Т-лимфоциты делят на Т-супрессоры и Т-хелперы (помощники), Т-контрсупрессоры. Т-супрессоры относятся к регуляторным клеткам, контролирующим процессы дифференцировки В-лимфоцитов и эффекторную активность Т-лимфоцитов. Они тормозят включение В-лимфоцитов
в пролиферацию и дифференцировку, угнетая тем самым выработку специфических антител, цитотоксические и другие иммунные реакции. Т-супрессорам принадлежит решающая роль в системе регуляции иммунного ответа. Одним из механизмов действия супрессоров являются распознавание идиотипических рецепторов Т- и В-лимфоцитов и угнетение их активности. Т-хелперы также выполняют иммунорегуляторные функции. Именно они, воздействуя на В-лимфоциты, обеспечивают развитие и накопление В-клеток, интенсивно продуцирующих антитела против иммунизирующего агента. Т-хелперы распознают последний и синтезируют более 20 разнообразных лим-фокинов, регулирующих иммунные реакции и помогающих В-клеткам, а также макрофагам и Т-киллерам. Разновидность Т-хелперов — индукторы, активируют Т-супрессоры. Т-хелперы играют вспомогательную роль в процессах антителообразо-вания, стимулируют пролиферацию и дифференцировку В-лимфоцитов в направлении иммуноглобулинсинтезирующих клеток при действии Т-зависимых антигенов. Общий эффект взаимодействий между Т-клетками после антигенной стимуляции состоит в восстановлении гомеостатического баланса на новом уровне с дифференцировкой специфических по отношению к антигенам кланов Т- и В-лимфоцитов. Контрсупрессоры — популяция Т-клеток, воздействующих на Т-супрессоры и блокирующих активность высвобождаемых ими медиаторов. Нарушение равновесия между физиологической функцией местных супрессоров и антисупрессорной регуляцией в организме способствует развитию хронических воспалительных заболеваний органов полости рта (например, болезней пародонта) и других отделов пищеварительного тракта.
Таким образом, для поддержания нормального иммунного ответа необходимо определенное соотношение Т-хелперных и Т-суп-рессорных лимфоцитов. Его нарушение приводит к развитию иммунопатологических процессов.
Гуморальную форму иммунитета, реализуемую посредством специфических антител, обеспечивают В-лимфоциты. Родоначальницей В-лимфоцитов является гемопоэтическая стволовая клетка. В-лимфоциты созревают в эмбриональной печени, затем в костном мозге, мигрируют в кровь и периферические лимфоидные органы, локализуясь в основном в лимфатических узлах и белой пульпе селезенки. В-лимфоциты содержатся также в тканях печени, пищеварительного тракта. Продолжительность их жизни колеблется от нескольких дней до нескольких недель, способность к рециркуляции выражена слабо. Основной маркер В-лимфоцитов — мембранный иммуноглобулин — представляет собой рецептор для антигена, обладающий высокой степенью специфичности. Иммуноглобулины встроены в мембраны В-лимфоцитов «хвостовой» частью
внутрь, а рецепторной — наружу. Последовательная трансформация В-лимфоцитов в плазматические клетки, продуцирующие антитела, активируется двумя сигналами — специфическим и неспецифическим. Неспецифический сигнал осуществляют медиаторы, синтезируемые макрофагами и Т-клетками индукторно-хелперного ряда. Эти медиаторы определяют деление и пролиферацию клеток. Специфический сигнал исходит от антигена и регулирует дифференцировку клеток. Средняя продолжительность жизни зрелых плазматических клеток слизистой оболочки кишечника, синтезирующих IgA, составляет около 5 сут. Синтез иммуноглобулинов осуществляется клетками плазмоцитарного ряда лимфатических узлов, костного мозга, селезенки, аппендикса. В результате контакта с антигеном В-лимфоциты могут трансформироваться не только в плазма-тические клетки, секретирующие иммуноглобулины, но и в долгоживущие клетки иммунной памяти.
Важнейшим функциональным свойством иммуноглобулинов является их способность идентифицировать антиген, активировать систему комплемента, фиксировать его отдельные компоненты, взаимодействовать с мембранами различных типов клеток. Иммуноглобулины могут проходить через эпителиальные мембраны, чем объясняется их присутствие в слюне, тканевой жидкости, внесосудистом пространстве, слизистой оболочке пищеварительного тракта.
Суть специфичности иммунных ферментов сводится к взаимодействию антигена и антитела, основанному на простран- ственном соответствии, комплементарности между ними. Ос- новное биологическое свойство иммуноглобулинов — связывание антигена и выделение его из организма.
Реакция связывания антителом антигена является универсальным механизмом элиминации антигена из внутренней среды организма. Иммунные комплексы, образующиеся при этом, поглощаются макрофагами печени, селезенки, нейтро-филами, подвергаются биологической трансформации и в норме удаляются из организма. При ряде заболеваний пищеварительного тракта, например при токсико-аллергических стоматитах, имеет место гипериммуноглобулинемия, вызванная стимуляцией антигенами — бактериями, токсинами, вирусами, другими инфекционными агентами, пищевыми белками, присутствующими в пищеварительном тракте, лекарственными препаратами. В физиологических условиях клетки интактной печени, прежде всего звездчатые эндотелиоциты, инактивируют бактерии, поступающие из воротной вены, защищая организм от сенсибилизирующего и токсического их действия. При на- рушении функциональной способности печени обезвреживание бактерий нередко снижается, антигенная активность последних растет. Кроме того, при нарушении циркуляции некото-
рые микроорганизмы кишечника или антигенные компоненты после абсорбции из пищеварительного тракта, минуя поврежденную печень, оказывают непосредственное воздействие на иммунокомпетентные клетки селезенки и лимфоидной ткани кишечника, вызывая поликлональную стимуляцию В-лимфо-цитов и образование антител. Гипериммуноглобулинемия может возникнуть при нарушении функциональной активности клеток-супрессоров и клеток-хелперов.
Антитела к антигенам пищевых продуктов. Хронические болезни органов пищеварения в ряде случаев сопровождаются появлением в циркулирующей крови антител, реагирующих с антигенами пищевых продуктов — чаще всего гликопротеида-ми с низкой молекулярной массой, содержащимися в молоке, яйцах, рыбе, цитрусовых. Большую роль в развитии аллергических реакций на пищевые вещества играет гистамин, обнаруженный в томатах, бобах, клубнике, какао, лесных орехах. Поступление экзогенного гистамина провоцирует аллергические реакции. Значительную роль в сенсибилизации к пищевым продуктам играют пищевые добавки, консерванты и ан-тиоксиданты — салицилаты, сульфаты, сульфиты. В состав коровьего молока входят более 20 компонентов, способных вызвать аллергические реакции немедленного и замедленного типов. Наиболее активным аллергеном является бета-лактоглобу-лин. В составе яичного белка выраженным аллергенным компонентом является овомукоид, обладающий антитрипсинной активностью, способствующей длительному сохранению его аллергенных свойств в кишечнике. Аллергены рыбных продуктов относятся к группе саркоплазматических белков, устойчивых к тепловой и ферментативной обработке. Основные аллергенные компоненты мяса содержатся как в собственно мышечной ткани, так и в сыворотке крови. Например, глобулин бычьей сыворотки сходен по химическому строению и иммунологическим свойствам с бета-лактоглобулином коровьего молока. Антитела к пищевым аллергенам относятся в основном в классам IgE, IgA, IgG и опосредуют реагиновый тип гиперчувствительности. Взаимодействие антител со специфическими пищевыми аллергенами в присутствии компонентов комплемента (анафилатоксинов) вызывает дегрануляцию тучных клеток тканевого функционального элемента, а также высвобождение гистамина базофилами. Аллергические антитела к пищевым продуктам принимают участие в патогенезе пищевой аллергии, протекающей по типу токсико-аллергичес-ких стоматитов, целиакии, болезни Крона, неспецифического язвенного колита, гиперэозинофильного гастроэнтерита.
Специфическое взаимодействие антигена с антителом сопровождается образованием иммунного комплекса. Этот физиологический процесс представляет собой механизм защиты
организма, способствующий элиминации антигенов. Иммунные комплексы взаимодействуют с комплементом и практически со всеми клетками крови — нейтрофилами, эозинофилами, лимфоцитами, тромбоцитами, а также с рецепторами клеток многих органов и тканей — эндотелия сосудов, клеток печени, клеток гломерулярного аппарата почек. Они стимулируют фагоцитоз, способствуя высвобождению содержимого лизосом, включая разрушающие ферменты, катионные белки, вазоак-тивные амины, медиаторы и другие биологически активные вещества. Фагоцитирующие клетки, в том числе и фиксиро- ванные тканевые макрофаги печени, селезенки и других органов, переваривают чужеродные вещества, участвуют в катаболизме иммунных комплексов, удаляют их из организма. Печень и селезенка являются основными эффекторными органами фагоцитарной мононуклеарной системы, захватывающей и элиминирующей циркулирующие иммунные комплексы, включающие прежде всего антигены кишечного происхождения. Кинетика иммунного ответа характеризуется серией взаимодействий и сменой взаимоотношений различных клеточных систем, регулирующих и осуществляющих эффекторную фазу иммунного ответа. Большая роль в механизмах межклеточного взаимодействия принадлежит клеточным медиаторам — лим- фокинам, монокинам, фиброкинам.
Медиаторы гиперчувствительности замедленного (лимфокины и монокины) и немедленного (гистамин, серотонин, кинины, лей- котриены) типов продуцируются различными клетками и характеризуются своими физико-химическими и биологическими свойствами. Лимфокины и ферменты относятся к пептидам; простагландины и лейкотриены — к липидам; гепарин — к гликозаминогликанам; гистамин — к аминам. Кинины отли- чаются сосудорасширяющим гистаминоподобным действием. Брадикинин способствует увеличению объема кровотока в ве-нулах. Медиаторы гиперчувствительности немедленного типа про-| дуцируются тучными клетками, базофилами, эозинофилами. Лейкотриены относятся к группе метаболитов арахидоновой кислоты, выделяемых лейкоцитами, оказывают сосудосужива- ющий эффект, влияют на проницаемость сосудистой стенки, с регулируют синтез гамма-интерферона, способствуют увеличе- нию его продукции. Лейкотриены играют большую роль в раз- витии анафилактического шока и аллергических реакций не- медленного типа. Будучи биологически активными соединени- ями, медиаторы взаимосвязаны между собой, функционируют по принципу обратной связи, дублирования, антагонизма и в совокупности составляют динамическую систему защиты организма. Для выработки лимфокинов необходимы не только сенсибилизированные антигенами Т-лимфоциты, но и их взаи-
функции.
Митогенный фактор играет важную роль в распознавании чужеродного антигена, способствует неспецифической клеточной пролиферации при иммунном ответе и его усилению.
Хемотаксические факторы изменяют хемотаксис, миграцию макрофагов, нейтрофилов, эозинофилов.
Действие факторов, угнетающих миграцию макрофагов и лейкоцитов, способствует накоплению фагоцитов в очаге иммунного воспаления, их активации.
Лимфотоксины опосредуют цитотоксическое действие на клетки-мишени, угнетают рост, вызывают деструкцию клеток. Активность лимфотоксических лимфоцитов-эффекторов и естественных клеток-киллеров в значительной мере зависит от интерлейкина-2, продуцируемого Т-хелперами.
Интерфероны подавляют репликацию вирусов, обладают некоторыми противоопухолевыми свойствами.
Ингибиторы синтеза ДНК подавляют иммунный ответ.
Фактор переноса способствует переходу несенсибилизирован-ных интактных лимфоцитов в состояние, эквивалентное сенсибилизации. Точкой приложения различных медиаторов служат лимфоциты, фагоциты, клетки-мишени. Ингибирующие лимфокины тормозят миграцию макрофагов и лейкоцитов в очаг воспаления, синтез ДНК, снижают темп деления клеток в культуре ткани, оказывают прямой цитотоксический эффект. Стимулирующие лимфокины ускоряют миграцию и пролиферацию клеток, усиливают положительный хемотаксис, увеличивают проницаемость клеточной мембраны. Не вызывает сомнения важная роль лимфокинов в регуляции процессов регенерации, репарации, гемопоэза, воспалительных процессов, неспецифической антимикробной защиты и специфического иммунного ответа.