

Вракин В.Ф, Сидорова М.В. |
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ |
|
|
довитости, повышенная эмбриональная смертность и появление уродств у потомства. Половые клетки плодов и новорожденных животных более чувствительны к повреждающим факторам, чем у взрослых животных.
В современном животноводстве все шире применяются различные методы стимуляции многоплодия и повышения оплодотворяемости. От одной коровы, стимулируя суперовуляцию, культивируя овулировавшие ооциты и пересаживая их другим коровам, можно в год получить до 60 телят (Ельчанинов, 1978). Однако на пути реализации этой заманчивой перспективы стоит много нерешенных вопросов. Дело в том, что при искусственно вызываемой полиовуляции (суперовуляции) нарушается синхронность между процессами роста, созревания и овуляции ооцитов. В яйцевод попадают половые клетки разной степени зрелости. Это приводит к большой гибели эмбрионов на ранних стадиях развития. У крупного рогатого скота при многоплодии часто появляются «интерсексы», особенно у молочных пород. Еще меньше изучены вопросы культивирования и пересадки яйцеклеток.
ОПЛОДОТВОРЕНИЕ И ЕГО БИОЛОГИЧЕСКОЕ ЗНАЧЕНИЕ
Оплодотворение — сложный комплекс физиологических процессов и реакций, приводящий к слиянию мужской и женской половых клеток в одну клетку — зиготу (zygotes — объединенный). В нем можно выделить осеменение и собственно оплодотворение.
Осеменение — это биологические процессы, обеспечивающие сближение гамет.
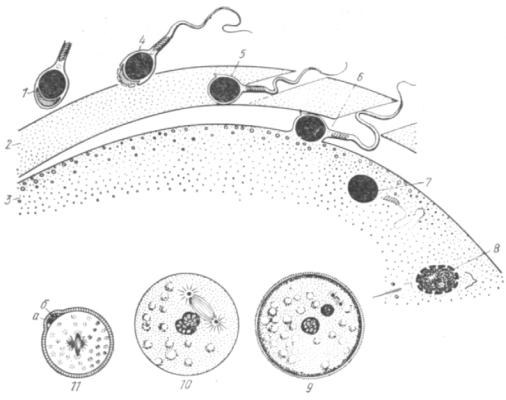
Собственно оплодотворение протекает в четыре стадии: 1) сближение половых клеток; 2) проникновение спермиев в блестящую оболочку; 3) проникновение в цитоплазму яйцеклетки; 4) слияние ядер половых клеток (рис. 18). В процессе оплодотворения две высокоспециализированные клетки — спер-мий и яйцеклетка — взаимодействуют друг с другом специфическим образом. Функция спермия — обеспечение встречи с яйцеклеткой, побуждение ее к развитию и внесение в нее отцовского наследственного материала. Функция яйцеклетки — обеспечение активации ядра спермия, привнесение материнского наследственного материала, накопление трофического, энергетического и структурного материала, а также регуляторных факторов (пока еще не выясненной природы), необходимых для начальных этапов развития зародыша.
У крупного и мелкого рогатого скота осеменение влагалищное, у лошадей и свиней — маточное. При этом во влагалище (у коровы, овцы и козы) или шейку матки (у кобылы и свиньи) попадают миллиарды спермиев (см. табл. 1). В половых путях самок спермин движутся благодаря реотаксису (способности двигаться против тока жидкости), а также присасывающего действия, производимого сокращением мускулатуры матки и яйцеводов. В результате через несколько часов (6—7 — у овец, 5—9—1 у коров) спермин оказываются в верхней трети яйцевода. Но доходит туда меньше 1 % спермиев. Основная их масса гибнет в пути: внедряются в слизистую оболочку по-
www.timacad.ru
Вракин В.Ф, Сидорова М.В. |
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ |
|
|
ловых путей, фагоцитируются лейкоцитами, склеиваются специальными веществами (фертилизинами), выделяемыми яйцеклеткой. Таким образом происходит как бы отбор спермиев, в процессе которого погибают атипичные и маложизненные клетки.
Рис. 18. Схема процесса оплодотворения:
1 —спермий; 2— блестящая оболочка; 3 — кортикальный слой цитоплазмы яйцеклетки; 4 — акросомная реакция; 5 — проникновение спермия через блестящую оболочку; 6 — слияние плазмолемм спермия и яйцеклетки; 7 — проникновение спермия в цитоплазму яйцеклетки; 8 — превращение головки спермия в мужской пронуклеус; 9 — сближение женского и мужского пронуклеусов; 10— слияние пронуклеусов; 11 — зигота (а и б —редукционные тельца).
Спермий с яйцеклеткой должны встретиться в верхней трети яйцевода. Если этого не произойдет и яйцеклетка до встречи со спермиями пройдет больший путь, она стареет и к концу яйцевода теряет способность к оплодотворению. Чтобы произошло оплодотворение, верхней трети яйцевода должно достигнуть по крайней мере несколько тысяч или десятков тысяч спермиев, так как эти клетки вступают в контакт с яйцеклеткой в результате случайного столкновения. Это следует учитывать при разбавлении спермы во время искусственного осеменения. Яйцеклетки выделяют вещества, ускоряющие движение и продлевающие период подвижности спермиев, но не выделяют специальных привлекающих веществ (хемотаксис отсутствует). Вероятность встречи увеличивается и тем, что половые клетки имеют разный электропотенциал: положительный у спермиев и отрицательный у яйцеклетки.
www.timacad.ru
Вракин В.Ф, Сидорова М.В. |
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ |
|
|
У большинства сельскохозяйственных животных лучистый венец яйцеклетки, попавшей в яйцевод, начинает разрушаться еще до встречи со спермиями. Однако под влиянием спермиев этот процесс совершается быстрее.
На первой стадии оплодотворения — при контакте яйцеклетки со спермиями в последних происходит акросомная реакция: частично разрушается цитолемма и наружная оболочка акросомы. Гиалуронидаза, заключенная в акросомах спермиев, изливается и разрушает остатки лучистого венца. Он рассыпается, блестящая оболочка яйцеклетки разрыхляется. Эта реакция неспецифическая, в ней могут участвовать и спермий других видов животных (что используется порой в животноводстве для экономии семени особо ценных производителей при искусственном осеменении).
Вторая стадия оплодотворения видоспецифична. В ней участвует несколько десятков спермиев своего вида. Они проходят сквозь прозрачную оболочку благодаря ферментам, связанным с внутренней мембраной акросомы. Приблизившись к поверхности яйцеклетки, один из спермиев сливается с ее цитолеммой боковой поверхностью головки — наступает третья стадия оплодотворения.
В месте контакта спермия и яйцеклетки частично расплавляются мембраны и цитоплазмы клеток сливаются. Мембрана спермия входит составной частью в мембрану яйцеклетки. Внутрь яйцеклетки спермий может войти целиком, а может только головкой, шейкой и средним отделом хвоста (у свиньи).
Слияние мембран спермия и яйцеклетки служит своего рода толчком для пробуждения яйца. С этого момента снимается блокада мейоза и он продолжается с той стадии, на которой остановился во время овуляции (завершается первое и начинается второе деление созревания). Резко активизируется метаболизм яйцеклетки. Развивается кортикальная реакция. Она состоит в том, что кортикальные гранулы подходят к цитолемме и их содержимое изливается в сторону прозрачной оболочки. Образуется перивителлиновое пространство, в результате чего яйцеклетка отслаивается от прозрачной (желточной) оболочки. Часть гранул входит в состав желточной оболочки, которая при этом утолщается и преобразуется в оболочку оплодотворения. Материал кортикальных гранул видоизменяет и цитолемму яйца. Она становится плотной и не проницаемой для других спермиев, так как покрывается гиалиновым слоем. Тем самым кортикальная реакция является механизмом, предохраняющим от проникновения сверхчисленных спермиев, и образует оболочку, под которой развивается зародыш до тех пор, пока он не покидает яйцовые оболочки.
Кортикальная реакция начинается от места слияния мембран спермия и яйцеклетки и распространяется отсюда по всей поверхности яйца. У олиголецитальных яиц это происходит быстро, в результате чего обеспечивается моноспермия — только один спермий попадает в яйцеклетку. У животных с полилецитальными яйцами (птицы, рептилии) наблюдается физиологическая полиспермия (кортикальная реакция у них развивается медленно и несколько спермиев могут за это время проникнуть внутрь яйца). Однако и в том и дру-
www.timacad.ru
Вракин В.Ф, Сидорова М.В. |
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ |
|
|
гом случае четвертая стадия оплодотворения (слияния ядер половых клеток) протекает аналогично. С ядром яйцеклетки сливается ядро лишь одного спермия.
Дальнейшая судьба частей спермия в цитоплазме яйцеклетки различна. Одни включаются в развитие, другие дегенерируют. Ядро спермия преобразуется в мужской пронуклеус (предъядрие) и проникает все глубже в цитоплазму, в нем за 20—30 мин реорганизуется и реактивируется хроматин, а его объем увеличивается в 50 раз. К моменту образования мужского пронуклеуса в ядре яйцеклетки завершается второе деление созревания, выделяется второе редукционное тельце и ядро преобразуется в женский пронуклеус. Пронуклеусы сближаются друг с другом. В это время в них происходит редупликация ДНК, а после ее завершения— профаза митоза. На основе центриолей центросомы формируется веретено деления. Сблизившиеся пронуклеусы сливаются. Конечная стадия их слияния — образование единой метафазной экваториальной пластинки уже не отдельных половых клеток, а зиготы. Оплодотворение завершилось — началось развитие нового организма.
Значение оплодотворения состоит в том, что оно приводит к слиянию отцовского и материнского геномов и таким образом является средством передачи наследственных свойств новому организму. В результате в стаде, породе, популяции возникает множество новых комбинаций наследственных факторов, создается многообразный генофонд, который служит материалом для отбора в ходе хозяйственной деятельности человека.
Клетка, появившаяся в результате оплодотворения—зигота, отличается от яйцеклетки своими биологическими, морфологическими и физикохимическими свойствами. Она обладает большой жизненностью, более широкой наследственной основой. Однако эти качества не абсолютны и во многом зависят как от возраста и состояния здоровья родителей, так и от сроков осеменения и оплодотворения. Близкородственное спаривание (инбридинг), как правило, приводит к появлению малочисленного слабого потомства. С возрастом матери увеличивается количество хромосомных нарушений. Несвоевременное осеменение приводит к «старению» половых клеток и, как следствие этого, к снижению их жизнеспособности и способности к оплодотворению.
Вопросы для самопроверки. 1. Опишите процессы, происходящие при сперматогенезе и оогенезе, и влияние на них внешних и внутренних факторов. 2. Какие черты сходства и различия сперматогенеза и оогенеза вы знаете? 3. В чем особенность стадии роста оогенеза? 4. Каково строение зрелого спермия и зрелой яйцеклетки? 5. Значение оплодотворения и его морфология.
www.timacad.ru
Вракин В.Ф, Сидорова М.В. |
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ |
|
|
Глава 5. РАННИЕ ЭТАПЫ ЭМБРИОНАЛЬНОГО РАЗВИТИЯ. ДРОБЛЕНИЕ
Сразу после образования зигота приступает к делению митозом. Деления быстро следуют одно за другим, митотический цикл клеток короткий, они не дорастают до размеров материнской, поэтому дочерние клетки с каждым делением становятся все мельче. Такой процесс называется дроблением, а клетки—бластомерами. Ядра бластомеров не уменьшаются, так как каждому делению предшествует редупликация ДНК. Ясно, что при каждом делении соотношение ядра и цитоплазмы в клетках изменяется. Дробление прекращается тогда, когда в бластомерах устанавливается ядерно-плазменное соотношение, типичное для соматических клеток данного вида животного. В клетках начинается активный белковый синтез, скорость деления снижается, каждая дочерняя клетка дорастает до размеров материнской, а размеры зародыша резко возрастают.
Скорость и характер дробления неодинаковы у разных животных и зависят от количества желтка в яйцеклетке. Олиголецитальные яйцеклетки (млекопитающие) привносят в зиготу мало желтка, он не мешает зиготе делиться полностью — от анимального до вегетативного полюса. Если при этом яйцеклетка была изолецитальная (ланцетник) —бластомеры получаются одинакового размера. Такое дробление называется полным равномерным. В телолецитальных яйцах бластомеры разных размеров, так как желток, скопившийся у вегетативного полюса, мешает делению и здесь образуются более крупные клетки, как, например, у амфибий. Это полное неравномерное дробление. Если яйца полилецитальные (рыбы, рептилии, птицы), крупные массы желтка настолько мешают дроблению, что делятся лишь клетки анимального полюса. Вся остальная масса остается в виде нераздробившегося желтка. Такой тип дробления называется частичным или дискоидальным (бластомеры в процессе дробления располагаются в форме диска).
В период дробления клетки делятся в определенном порядке. Первая и вторая борозды дробления идут от анимального к вегетативному полюсу и называются меридианальными. Третья борозда — широтная. Дальше идут чередования меридианальных, широтных и тангенциальных (параллельно поверхности) борозд, в результате чего правильность расположения бластомеров нарушается. Дольше всего правильность их расположения сохраняется при полном равномерном дроблении. Быстрее всего она нарушается при частичном дроблении (цв. табл. I) уже после 2—3-го деления.
Бластомеры во время дробления тесно связаны друг с другом, образуется как бы плотный комок из клеток — морила, а затем пузырек с полостью — бластула. Стенка бластулы называется бластодерма, а полость — бластоцель. Клетки бластодермы, расположенные на месте бывшего анимального полюса, образуют крышу, а расположенные на месте вегетативного полюса
— дно бластулы. Между ними находится краевая зона. При полном равномерном дроблении изоолиголецитальных яиц образуется бластула с бластодермой из одного слоя почти равных клеток и большим бластоцелем. Это це-
www.timacad.ru