

2. Энергетический обмен и его роль в явлениях фитоиммунитета. Окисление, окислительное фосфорилирование.
Для отражения атаки проникшего в клетку паразита необходима энергия, она нужна для образования защитных веществ, новых клеток и тканей, а также для детоксикации метаболитов паразита. Основным источником энергии является дыхание.
Из общей суммарной формулы дыхания следует, что при окислении кислородом одной молекулы глюкозы выделяется энергия:
C6H12О6
+ 6О2
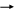 6CО2
+ 6H2О
+ 674 ккал (2824 кДж)
6CО2
+ 6H2О
+ 674 ккал (2824 кДж)
Необходимый кислород клетка получает из воздуха. Однако одноразовое выделение большого количества энергии может оказать разрушающее действие на живую клетку. Поэтому высвобождение энергии происходит в несколько этапов с запасанием ее в АТФ. Процесс запасания энергии является многоступенчатым и называется окислительным фосфорилированием.
Окислительное фосфорилирование очень чувствительно к различным воздействиям, причем чувствительность фосфорилирования во много раз больше сопряженного с ним окисления. В случае неблагоприятного воздействия поглощение неорганического фосфора может полностью прекращаться, в то время как окисление продолжается и даже становится интенсивнее. Внешняя картина дыхания при этом не меняется или поглощение кислорода тканью даже усиливается. Такое дыхание уже не является биологически полноценным и называется разобщенным, а вещества, вызывающие его,— разобщителями, или разобщающими ядами.
Живая система способна самостоятельно регулировать степень сопряжения и фосфорилирования.
Было сделано немало попыток связать устойчивость растений к инфекционным болезням с уровнем дыхания их тканей. На первый взгляд такой подход вполне обоснован, так как дыхание является одним из самых точных показателей жизнедеятельности любого организма.
В ответ на инфекцию возрастает интенсивность дыхания и повышается температура тканей. Пораженная ткань может выделять в 10 раз больше углекислого газа, чем интактная. Повышение интенсивности обмена под влиянием паразита происходит не только в зараженных, но и в здоровых соседних тканях. Следовательно, под влиянием местного заражения происходит повышение интенсивности обмена веществ всего плода, хотя по мере удаления от очага инфекции степень реагирования тканей сильно понижается.
Вначале предполагалось, что более высокой устойчивостью должны обладать растения, с низким уровнем дыхательного газообмена вследствие того, что с усилением дыхания увеличивается расход дыхательного материала, а это должно привести к более быстрому истощению растительных тканей.
Другая диаметрально противоположная точка зрения о роли дыхания в устойчивости картофеля при хранении состоит в том, что наиболее лежкими являются те клубни, которые характеризуются высоким уровнем дыхания.
Не меньшее число данных свидетельствует и об отсутствии прямой связи между лежкостью клубней и присущим их тканям уровнем дыхательного газообмена.
Многочисленные исследования позволяют утверждать, что на основании определения интенсивности дыхания здоровых клубней практически невозможно судить об их лежкости.
Более определенные данные были получены при изучении интенсивности дыхания пораженных клубней. Замечено, что при заражении растительных тканей паразитарными грибами, бактериями, вирусами, фитогельминтами их дыхание заметно усиливается. Если растение успешно справляется с поражением, то дыхание возвращается к норме. Спад дыхания наблюдается и при заболевании растения, но уже по причине отмирания клеток.
При поражении фитопатогенными микроорганизмами активность дыхания у устойчивых к ним растений значительно выше, чем у восприимчивых.
Дело не только в усилении или ослаблении интенсивности дыхания. Например, облучение картофеля вызывает подъем дыхания, но одновременно нарушается сопряженность между окислением и фосфорилированием. В результате образующаяся при дыхании энергия рассеивается в виде тепла и не может быть использована на синтез веществ, необходимых для роста, а также для защиты от инфекции.
Следовательно, необходимо изучать энергетическую эффективность дыхательного газообмена и на что используется энергия, на какие процессы и вещества, которые, в конечном счете, определяют устойчивость к фитопатогенным микроорганизмам или состояние покоя и переход к активному росту.
Пример: В ответ на механическое поранение и внедрение гриба возбудителя фитофтороза в клубнях картофеля усиливаются дыхание, активность полифенолоксидазы и пероксидазы, окислительное фосфорилирование, образуются новые митохондрии. Однако в результате этих реакций, однотипных как для механического поранения, так и заражения несовместимой расой фитофторы образуются совершенно разные вещества.
При всей очевидности связи дыхания с жизнью растения механизм зависимости того или иного проявления жизнедеятельности от дыхания удается понять лишь в результате углубленного физиологического анализа дыхательного процесса и связанных с ним превращений веществ и энергии.
Только при условии сопряженности процессов окисления и фосфорилирования может быть наиболее полно использована устойчивость, которая присуща растительным тканям.