

50. Обмен аминокислот в тканях.
Промежуточный обмен аминокислот делят на общие пути обмена и индивидуальные превращения отдельных аминокислот. Распад аминокислот осуществляется по гораздо более сложным механизмам, чем распад жирных кислот или моносахаридов, что объясняется их структурным разнообразием.
Общие пути превращения аминокислот включают реакции дезаминирования, трансаминирования, декарбоксилирования, биосинтеза.
Все аминокислоты в конечном итоге распадаются до ПВК, либо до ацетил–КоА, либо до промежуточных продуктов ЦТК. Азот, входящий в состав всех аминокислот, но не входящий в состав конечных продуктов их распада, отщепляется на самых первых этапах в виде NН3, превращается в мочевину и выводится из организма.
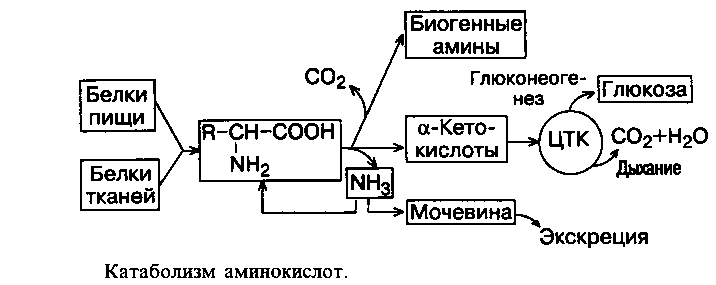
Дезаминирование – отщепление от аминокислоты аминной группы –NН2 при участии ферментов дезаминаз или оксидаз, сопровождающееся выделением энергии.
Во всех случаях NН2- группа аминокислоты освобождается в виде аммиака.
Существует 4 типа дезаминирования аминокислот.
-
Восстановительное дезаминирование:
![]()
-
Гидролитическое дезаминирование:
![]()
Эти способы характерны для бактерий.
3.Внутримолекулярное дезаминирование:
![]()
Данный вид дезаминирования характерен для растений, бактерий, в животном организме так дезаминируется гистидин.
4.Окислительное дезаминирование:
![]() Является
преобладающим типом дезаминирования
в организме животных. Происходит в
клетках печени и почек. Помимо аммиака,
продуктами дезаминирования являются
жирные кислоты, оксикислоты и кетокислоты.
Механизм
окислительного дезаминирования
аминокислот, протекающего в две стадии:
Является
преобладающим типом дезаминирования
в организме животных. Происходит в
клетках печени и почек. Помимо аммиака,
продуктами дезаминирования являются
жирные кислоты, оксикислоты и кетокислоты.
Механизм
окислительного дезаминирования
аминокислот, протекающего в две стадии:
![]()
Первая стадия является ферментативной (при участии дегидрогеназ) и завершается образованием неустойчивого промежуточного продукта (иминокислота), который на второй стадии в присутствии воды распадается на аммиак и - кетокислоту.
Наиболее важной в окислительном дезаминировании является глутаматдегидрогеназная реакция:

Трансаминирование Большинство аминокислот теряют аминогруппу в результате реакций трансаминирования.
Трансаминирование (переаминирование) – межмолекулярный перенос аминогруппы с -аминокислоты на -кетокислоту без промежуточного образования аммиака.
![]()
Впервые реакции трансаминирования были открыты в 1937 г. при изучении дезаминирования глутаминовой кислоты в мышечной ткани. Было замечено, что при добавлении к гомогенату мышц АЛА и - кетоглутаровой кислоты приводило к образованию соответственно ПВК и глутаминовой кислот.
Реакции трансаминирования являются обратимыми и универсальными для всех живых организмов; протекают при участии специфических ферментов аминотрансфераз. Наиболее распространенными из них являются: глутаматаминотрансфераза (переносит аминогруппу с любой аминокислоты на -кетоглутаровую кислоту, которая при этом превращается в глутаминовую); аспартатаминотрансфераза (переносит аминогруппу с любой аминокислоты на ЩУК с образованием аспарагиновой кислоты); аланинаминотрансфераза (переносит аминогруппу с любой аминокислоты на ПВК с образованием аланина).
Все аминотрансферазы в качестве кофермента содержат пиридоксальфосфат (производное вит. В6). Каждая аминотрансфераза специфична к определенным парам -амино- и -кетокислот. В организме человека имеется более десятка аминотрансфераз. Замещение кетогруппы в -кетокислоте на аминогруппу представляет собой конечную стадию синтеза аминокислоты. Наоборот, замещение NН2-группы в аминокислоте на кетогруппу − первая стадия катаболизма аминокислот. Следовательно, трансаминирование может служить как для синтеза, так и для катаболизма аминокислот. Аминотрансферазы содержатся практически во всех органах, но наиболее активно реакции трансаминирования протекают в печени. Функциональное значение трансаминирования в разных органах различно. Например, работающая мышца выделяет в кровь наряду с молочной кислотой значительные количества количества аланина. АЛА образуется в мышце из ПВК путем трансаминирования. Из кровотока АЛА поглощается печенью, где в результате трансаминирования вновь превращается в ПВК, а ПВК используется для глюконеогенеаза (глюкозо-аланиновый цикл). В результате реакций трансаминирования общее количество аминокислот аминокислот в организме не изменяется, т.к. в каждой реакции одна аминокислота превращается в безазотистый остаток (в -кетокислоту), а один безазотистый остаток – в новую аминокислоту. Напротив, дезаминирование ведет к уменьшению общего количества аминокислот, т.к. NН2-группа не используется для образования новой аминокислоты, а превращается в NН3.
Клиническое значение определения активности трансаминаз. Широкое распространение и высокая активность трансаминаз в органах и тканях человека, а также сравнительно низкие величины активности этих ферментов в крови послужили основанием для определения уровня ряда трансаминаз в сыворотке крови человека при органических и функциональных поражениях разных органов. Для клинических целей наибольшее значение имеют две трансаминазы - аспартатаминотрансфераза (AcAT) и аланинаминотрансфераза (АлАТ).
Повышение уровня трансаминаз в сыворотке крови отмечено при инфаркте миокарда, при некоторых заболеваниях мышц, в частности при обширных травмах, гангрене конечностей и прогрессивной мышечной дистрофии, поражениях клеток печени, например при гепатитах.
Непрямое дезаминирование Является общим для обмена всех аминокислот, кроме глутаминовой кислоты (т.к. ГЛУ подвергается прямому окислительному дезаминированию). Образующаяся в ходе трансаминирования глутаминовая кислота способна к непосредственному дезаминированию, т.е. к отщеплению азота в виде аммиака. Непрямое дезаминирование происходит в два этапа:
1. Трансаминирование – аминокислота передает аминогруппу -КГ и при этом превращается в кетоформу, а -КГ – в глутаминовую:
![]() 2.
Окислительное
дезаминирование ГЛУ с выделением
аммиака.
2.
Окислительное
дезаминирование ГЛУ с выделением
аммиака.
![]() -кетоглутаровая
кислота может вновь вступать в реакции
трансаминирования с другой аминокислотой,
затем дезаминироваться. Поскольку обе
реакции (трансаминирование и дезаминирование
глутаминовой кислоты) являются обратимыми,
создаются условия для синтеза любой
аминокислоты, если в организме имеются
соответствующие -кетокислоты.
Организм человека и животных не наделен
способностью синтеза незаменимых
аминокислот, из-за отсутствия
соответствующих -кетокислот.
После дезаминирования углеродный скелет
аминокислоты либо окисляется в аэробных
условиях и служит источником энергии,
либо используется в биосинтезе соединений.
-кетоглутаровая
кислота может вновь вступать в реакции
трансаминирования с другой аминокислотой,
затем дезаминироваться. Поскольку обе
реакции (трансаминирование и дезаминирование
глутаминовой кислоты) являются обратимыми,
создаются условия для синтеза любой
аминокислоты, если в организме имеются
соответствующие -кетокислоты.
Организм человека и животных не наделен
способностью синтеза незаменимых
аминокислот, из-за отсутствия
соответствующих -кетокислот.
После дезаминирования углеродный скелет
аминокислоты либо окисляется в аэробных
условиях и служит источником энергии,
либо используется в биосинтезе соединений.
Декарбоксилирование аминокислот
Декарбоксилирование – процесс отщепления карбоксильной группы аминокислот в виде СО2. В живых организмах открыты 4 типа декарбоксилирования аминокислот. Для тканей животных характерно -декарбоксилирование, при котором от аминокислот отщепляется карбоксильная группа, расположенная по соседству с -углеродным атомом:
![]()
Продуктами реакции являются СО2 и биогенные амины. Реакции декарбоксилирования в отличие от других процессов промежуточного обмена аминокислот являются необратимыми. Они катализируются специфическими ферментами – декарбоксилазами аминокислот, простетическая группа которых представлена пиридоксальфосфатом, как и у аминотрансфераз. Таким образом, в двух совершенно различных процессах обмена участвует один и тот же кофермент. Несмотря на ограниченный круг аминокислот, подвергающихся декарбоксилированию в животных тканях, образующиеся продукты реакции – биогенные амины (гистамин, -аминомасляная кислота и др.) оказывают сильное фармакологическое действие на физиологические функции организма. Гистамин образуется при декарбоксилировании гистидина, оказывает широкий спектр биологического действия: вызывает расширение капилляров и повышение их проницаемости (обладает сосудорасширяющим действием в отличие от других биогенных аминов),понижает АД, стимулирует секрецию желудочного сока и слюны; сокращает гладкие мышцы легких, что может вызвать «гистаминовый шок», который проявляется как приступ удушья; участвует в развитии болевых ощущений. Большое количество гистамина образуется в очаге воспаления, что имеет определенный биологический смысл, вызывая расширение сосудов в очаге воспаления, гистамин тем самым ускоряет приток лейкоцитов, способствуя активации защитных сил организма. При повышенной чувствительности к гистамину в клинике используют антигистаминные препараты, оказывая влияние на рецепторы сосудов. -аминомасляная кислота (ГАМК) образуется при декарбоксилировании глутаминовой кислоты, оказывает тормозящее действие на ЦНС (нейрогуморальный ингибитор). Обнаружена в сером веществе головного мозга, ее введение в организм вызывает торможение в коре (центральное торможение). Серотонин образуется из триптофана в нейронах гипоталамуса, функционирует как нейромедиатор в ЦНС, оказывает мощное сосудосуживающее действие, регулирует АД, температуру тела, дыхание, почечную фильтрацию. Этаноламин образуется при декарбоксилировании серина. Используется для синтеза холина, ацетилхолина, фосфолипидов (фосфатидилэтаноламина, фосфатидилхолина). Дофамин образуется из тирозина в почках, надпочечниках, синаптических ганглиях и нервах, является нейромедиатором ингибирующего типа. В других клетках является предшественником других катехоламинов (адреналина и норадреналина). Норадреналин образуется в результате гидроксилирования дофамина в клетках нервной ткани, мозговом веществе надпочечников. Функционирует как медиатор. Адреналин − продукт метилирования норадреналина в клетках мозгового вещества надпочечников. Является гормоном. Накопление биогенных аминов может отрицательно сказываться на физиологическом статусе организма. Инактивация биогенных аминов происходит путем их дезаминирования и окисления под действием ФАД-зависимой моноаминооксидазы (МАО) в митохондриях и диаминооксидазы (ДАО) в цитозоле. Изменение концентрации биогенных аминов является причиной ряда патологических состояний. Например, при болезни Паркинсона наблюдается уменьшение количества дофамина и одним из способов лечения является снижение скорости инактивации дофамина под влиянием веществ − ингибиторов МАО.