

Nikolls_-_Ot_neyrona_k_mozgu
.pdfПредисловие редакторов русского перевода
Данная книга является переводом четвертого издания знаменитой и ставшей классической книги «От нейрона к мозгу». Первое издание этой книги, написанной Стефеном Куффлером и Джоном Николлсом в 1975 году, было переведено на русский язык и вышло в СССР в 1979 году под редакцией профессора Л. Г. Магазаника. Несмотря на большой тираж, книга быстро исчезла с прилавков магазинов и стала настольным учебником для нескольких поколений нейрофизиологов. Последующие английские издания книги были существенно переработаны новым коллективом авторов, однако ни второе, ни третье издание в России не выходили. Публикация на русском языке данного, четвертого издания стала возможной благодаря поддержке Российского фонда фундаментальных исследований. Возвращение к этой книге в начале нового тысячелетия кажется нам символичным — новое издание для нового поколения означает, надеемся, возрождение интереса к науке. Цель нового издания, декларированная в предисловии к английскому изданию, осталась той же, что и первого, написанного более четверти века назад — «описать способы передачи сигналов нервными клетками, как сигналы анализируются и как на основе этой интеграции возникают высшие функции мозга». Однако, фактически, это новая книга, написанная в соответствии с бурным развитием нейробиологии последних лет. В предисловии, специально написанном к русскому переводу четвертого издания, Джон Николлс особо выделяет предмет «нейробиология» как междисциплинарную науку, способную интегрировать знания и подходы смежных наук. В России нейробиология пока не выделена в отдельный предмет, как это произошло в большинстве известных западных университетов. Книга «От нейрона к мозгу» может стать для студентов, по сути дела, учебником по нейробиологии. Мы намеренно представили часть текста в подписях к рисункам как на русском, так и на оригинальном английском языке. Нам кажется, что это позволит студентам и начинающим ученым легче освоить специфическую терминологию и быстрее интегрироваться в интернациональное научное сообщество.
Новая книга сохранила узнаваемые черты первого издания, такие как доступность и простота толкования самых сложных явлений в сочетании с высокой научностью, логично отслеженная взаимосвязь различных вопросов нейробиологии, огромное количество ярких наглядных иллюстраций. Объяснение каждого процесса ведется логично и последовательно, мысль авторов четка и ясна. Приятной особенностью книги является то, что авторы не опускают спорные вопросы, описывают альтернативные точки зрения и указывают на нерешенные в настоящее время проблемы. Последнее особенно ценно, поскольку это указания экспертов в этой области на перспективные направления возможного дальнейшего развития нейронауки. Не оставлен без внимания и исторический анализ развития представлений о работе мозга, что делает книгу эмоциональной и увлекательной.
Постоянным автором и главным инициатором всех четырех изданий является Джон Николлс (John Nicolls), профессор Международной высшей школы (SISSA) в Триесте (Италия). Другими авторами являются известные нейробиологи Брюс Валлас (Bruce G.Wallace) и Роберт Мартин (A. Robert Martin) (University of Colorado), а также Пол Фукс (Paul A. Fuchs) (The John Hopkins University). Все авторы внесли существенный личный вклад в разработку излагаемых проблем, что обеспечило «взгляд изнутри» на многие проблемы нейробиологии и привнесло в изложение особую достоверность и точность.
Первое издание «От нейрона к мозгу» до сих пор входит как рекомендованная литература практически во все курсы, касающиеся работы мозга, для студентов медицинских и биологических вузов России. Надеемся, что новое, полностью переработанное современное издание займет такое же место.
П. Балабан Р. Гиниатуллин
Предисловие авторов к русскому изданию
При написании «От нейрона к мозгу» я и мои коллеги прежде всего преследовали цель создания легко читаемой книги, которая помогла бы студентам медицинских и биологических факультетов в освоении знаний о нервной системе. Мы надеялись, что книга будет полезна и исследователям в этой области, равно как и физикам, инженерам и молекулярным биологам.
Термины «нейробиология» и «нейронауки» вошли в обиход в 60-е годы XX в., когда Стивен Куффлер создал в медицинской школе Гарвардского университета первый факультет, сотрудниками которого стали физиологи, анатомы и биохимики. Работая вместе, они решали проблемы функционирования и развития нервной системы, исследовали молекулярные механизмы работы мозга. До этого ученые этих специальностей работали отдельно, и учебники того времени отражают существовавшее разделение. Настоящая книга представляет собой особый подход к экспериментальному анализу и отражает развитие ключевых концепций науки о мозге вне зависимости от используемых методов.
Авторы книги очень надеялись на то, что богато иллюстрированный материал будет легко доступен каждому студенту, который просто пролистает книгу и решит, стоит ли ее прочесть. Несмотря на то, что английский язык сейчас стал необходимой частью образования исследователя, текст не на родном языке всегда воспринимается труднее. Теряется та часть научного подхода, которую мы старались передать в книге: красота науки, элегантность экспериментов, связь между классическими исследованиями прошлых лет и открывающиеся сегодня горизонты будущего. Именно поэтому известие о том, что вскоре появится русский перевод последнего издания нашей книги, так радует авторов. Стивен Куффлер всегда надеялся, что его книга будет переведена на другие языки. Первое издание книги было блестяще переведено в 1979 году нашим другом профессором Львом Магазаником из Института эволюционной физиологии и биохимии АН СССР (Ленинград). С тех пор было сделано столько ключевых открытий и настолько изменилось наше понимание механизмов работы мозга, что появление на русском языке книги для студентов о современной нейронауке стало необходимостью.
Многие известные ученые России и бывшего СССР внесли огромный вклад в наше понимание биофизических, нейрохимических и высших функций мозга, что послужило основой для развития исследований синаптической передачи, передачи зрительного сигнала, механизмов моторного контроля, функций коры мозга и механизмов обучения. Этот вклад и создание признанных научных школ также лежат в основе появившегося в последнее время в мире нового интереса к нейронаукам.
Как всегда, авторы благодарят нашего издателя Э. Синауэра за помощь в издании оригинала и русского перевода. Мы благодарим наших друзей профессоров П. Балабана и Р. Гиниатуллина за их усилия по редактированию и выпуску российского издания. Переводчики Л. Хируг, Р. Хасипов и А. Галкин, вместе с редакторами быстро и профессионально завершили подготовку издания. Несомненным достоинством книги является сохранение оригинальных подписей на английском языке, что позволяет легче перейти к анализу мировой литературы.
Мы надеемся, что настоящее издание послужит дальнейшим стимулом к развитию интереса к бурно развивающимся в настоящее время нейронаукам, вклад в которые российских ученых неоспорим.
Джон Николлс 2003 г., Триест, Италия

Раздел I. ВВЕДЕНИЕ
Глава 1. Принципы передачи информации и структурная организация мозга
Центральная нервная система представляет собой непрерывно работающий конгломерат клеток, которые постоянно получают информацию, анализируют ее, перерабатывают и принимают решения. Мозг способен также брать инициативу на себя и производить координированные, эффективные мышечные сокращения для ходьбы, глотания или пения. Для регуляции многих аспектов поведения и для прямого или непрямого контроля всего тела, нервная система обладает огромным количеством линий коммуникаций, обеспечиваемых нервными клетками (нейронами). Нейроны представляют собой основную единицу, или составной блок, мозга. Наша задача — изучить значение их взаимодействия.
Одна из целей этой книги приведена в названии. В последующих главах мы попытаемся объяснить поведение и сложные функции мозга в терминах активности нервных клеток. Второй целью является рассмотрение клеточных и молекулярных механизмов межнейронного взаимодействия. Третья цель связана с изучением способов образования структур и связей, которые лежат в основе функций развивающегося организма, как эти функции и связи изменяются с опытом и могут восстановиться после повреждения. В настоящей главе мы суммируем ключевые концепции и основы рассматриваемых явлений.
Взаимосвязи в простых нервных системах
События, которые происходят при реализации простых рефлексов, могут быть прослежены и проанализированы детально. Например, когда по коленной связке ударяют маленьким молоточком, мышцы и сухожилия бедра растягиваются и электрические импульсы по сенсорным нервным волокнам идут в спинной мозг, в котором возбуждаются моторные клетки, производя импульсы и активируя мышечные сокращения. Конечным результатом является распрямление ноги в коленном суставе. Такие упрощенные схемы очень важны для регулировки мышечных сокращений, управляющих движениями конечностей. В таком простом рефлексе, в котором стимул ведет к определенному выходу, роль сигналов и взаимодействий всего двух видов клеток может быть успешно проанализирована.
Сложные нейронные сети и высшие функции мозга
Анализ взаимодействия нейронов в сложных путях, вовлекающих в буквальном смысле миллионы нейронов, существенно более труден, чем анализ простых рефлексов. Пере-
16 |
Раздел I. Введение |
дача информации в мозг при восприятии звука, прикосновения, запаха или зрительного образа требует последовательного вовлечения нейрона за нейроном, так же как и при выполнении простого произвольного движения. Серьезная проблема при анализе взаимодействия нейронов и структуры сети возникает из-за плотной упаковки нервных клеток, сложности их взаимосвязей и обилия типов клеток. Мозг устроен не так, как печень, которая состоит из одинаковых популяций клеток. Если вы обнаружили, как работает одна область печени, то вы знаете очень много о печени в целом. Знания о мозжечке, однако, ничего не скажут вам о работе сетчатки или любой другой части центральной нервной системы.
Несмотря на огромную сложность нервной системы, сейчас возможно проанализировать много способов взаимодействия нейронов при восприятии. Например, записывая активность нейронов в пути от глаза к мозгу, можно проследить сигналы сначала в клетках, специфически отвечающих на свет, и затем, шаг за шагом, по последовательным переключениям, до высших центров мозга.
Интересной особенностью работы зрительной системы является способность выделять контрастные образы, цвета и движения в огромном диапазоне интенсивностей цвета. Когда вы читаете эту страницу, сигналы внутри глаза обеспечивают возможность для черных букв выделяться на белой странице в слабоосвещенной комнате или при ярком солнечном освещении Специфические связи в мозге образуют единую картину, несмотря на то, что два глаза расположены раздельно и сканируют отличающиеся области внешнего мира. Более того, существуют механизмы, обеспечивающие постоянство образа (хотя наши глаза непрерывно двигаются) и дающие точную информацию о расстоянии до страницы.
Каким образом связи нервных клеток обеспечивают подобные явления? Несмотря на то, что мы еще не способны дать полное объяснение, сейчас многое известно о том, как эти свойства зрения обеспечиваются простыми нейрональными сетями в глазе и на начальных стадиях переключения в мозге. Конечно, остается много вопросов о том, каковы связи между свойствами нейронов и поведением. Так, для того чтобы прочесть страницу, вы должны сохранять определенное
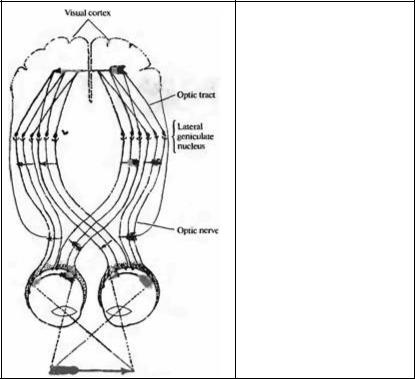
Рис. 1.1. Пути от глаза до мозга через оптический нерв и оптический тракт.
Fig. 1.1. Pathways from the Eyes to the Brain through the optic nerve and the optic tract. The interposed relay is the lateral geniculate nucleus. Arrows indicate how images are reversed by the lens and how the specific crossing of axons causes the right visual field to be represented in the left brain, and vice versa. The figure has been modified from an original by Ramo y CajaL which dates from 1892 (119091911] Eng. trans. 1995).
положение тела, головы и рук. Далее, мозг должен обеспечить постоянное увлажнение глазного яблока, постоянство дыхания и многие другие непроизвольные и неподконтрольные сознанию функции. Подобные проблемы, предусматривающие описание целостной картины координированных движений тела, выходят за рамки этой книги.
В дальнейшем мы рассмотрим принципы организации нервной клетки, возникновения и распространения электрических сигналов от нейрона к нейрону. Функционирование сетчатки является хорошим примером основных принципов работы нервной системы.
Глава 1. Передача информации и структурная организация мозга |
17 |
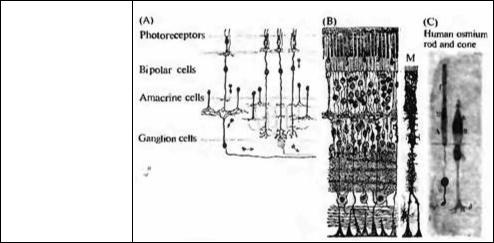
Рис. 1.2. Структура и связи клеток в сетчатке млекопитающих. (А) Схема направления сигнала от рецептора к оптическому нерву по Рамон-и-Кахалю. (В) Распределение по Рамон-и- Кахалю клеточных элементов сетчатки. (С) Рисунки палочки и колбочки сетчатки человека.
Fig. 1.2. Structure and Connections of Cells in the Mammalian Retina. The photoreceptors (rods and cones) connect to bipolar cells. Bipolar cells in turn connect to ganglion cells, whose axons constitute the optic nerve. Horizontal cells (not shown) and amacrine cells make connections that are predominantly horizontal. (A) The scheme proposed by Ramon y Cajal for the direction taken by signals as they pass from receptors to the optic nerve fibers. This scheme still holds in general but essential new pathways and feedback groups have been discovered since Ramon y Cajal's time.
(B) Ramon y Cajal's depiction of the cellular elements of the retina and their orderly arrangement. The Mueller cell (M) shown on the right is a satellite glial cell. (C) Drawings of a human rod (left) and cone (right) isolated from the retina. Light passes through the retina (in these drawings from bottom to top) to be absorbed by the outer segment (top) of the photoreceptor. There it produces a signal that spreads to the terminal to influence the next cell in line. By recording electrically from each cell in the retinal circuit we can follow signals step by step and understand how the meaning of the signals changes. (After Ramon y Cajal, 1995.)
§ 1. Строение сетчатки
Анализ зрительного мира зависит от информации, поступающей от сетчатки, где происходит первая стадия обработки, устанавливающая пределы для нашего восприятия. На рис. 1.1 показаны пути от глаза до высших центров мозга. Изображение, попадающее на сетчатку, перевернуто, но во всех других аспектах представляет собой добросовестное представление о внешнем мире. Каким образом эта картинка может быть передана в наш мозг посредством электрических сигналов, которые возникают в сетчатке и затем путешествуют по оптическим нервам?
Образы и связи нейронов
На рис. 1.2 показаны разные типы клеток и их расположение в сетчатке. Свет, попадающий в глаз, проходит сквозь слои прозрачных клеток и достигает фоторецепторов. Сигналы, передаваемые из глаза по волокнам оптического нерва, являются единственными информационными сигналами, на которых основано наше зрение.
Схема прохождения информации по сетчатке (рис. 1.2А) была предложена Сантьяго Рамон-и- Кахалем1) в конце XIX века. Он был одним из величайших исследователей нервной системы и проводил эксперименты на самых разных животных. Он сделал существенное обобщение о том, что форма и расположение нейронов, так же как область возникновения и конечная мишень нейрональных сигналов в сети, дают важнейшую информацию об функционировании нервной системы.
На рис. 1.2 ясно видно, что клетки в сетчатке, как и в других частях центральной нервной системы (ЦНС), очень плотно упакованы. Вначале морфологам приходилось разрывать нервную ткань на части, чтобы увидеть отдельные нервные клетки. Методы, при которых окрашивают все нейроны, практически бесполезны для исследования формы и связи клеток, потому что такие структуры, как сетчатка, выглядят подобно темному пятну переплетенных клеток и отростков. Электронная микрофотография на рис. 1.3 показывает, что экстраклеточное пространство вокруг нейронов и поддерживающих клеток составляет всего 25 нанометров в ширину. Большая часть рисунков Рамон-и-Кахаля была сделана с помощью метода окраски по Гольджи, ко-

18 |
Раздел I. Введение |
Рис. 1.3. Плотная упаковка нейронов в сетчатке обезьяны. Помечена одна палочка (R) и одна колбочка (С).
Fig. 1.3. Dense Packing of Neurons in Monkey (Macaque) Retina. This electron micrograph shows a characteristic feature of the central nervous system: The cell membranes are separated by narrow, fluid-filled clefts. The photoreceptors and their processes can be followed to the outer plexiform layer where their terminals contact bipolar and horizontal cells. One cone (C) and one rod (R) are labeled. (Micrograph kindly provided by P. Sterling and Y. Tsukamoto.)
торый окрашивает с помощью неизвестного механизма всего несколько случайных нейронов из всей популяции, но эти несколько нейронов окрашены полностью.
Схема на рис. 1.2 показывает принцип упорядоченного расположения нейронов в сетчатке. Легко отличить фоторецепторы, биполярные и ганглиозные клетки. Направление передачи идет от входа к выходу, от фоторецепторов к ганглиозным клеткам. Кроме того, два других типа клеток, горизонтальные и амакриновые, образуют связи, соединяющие разные пути. Одной из целей нейробиологии, присутствующей в рисунках Рамон-и-Кахаля, является стремление понять, как каждая клетка участвует в создании картины мира, которую мы наблюдаем.
Тело клетки, дендриты, аксоны
Ганглиозная клетка, показанная на рис. 1.4, иллюстрирует особенности строения нервных клеток, присущие всем нейронам центральной и периферической нервной системы. Клеточное тело содержит ядро и другие внутриклеточные органеллы, общие для всех клеток. Длинный отросток, который покидает тело клетки и образует связь с клеткой-- мишенью, называется аксоном. Термины дендрит, тело клетки и аксон применяются к отросткам, на которых входящие волокна образуют контакты, играющие роль принимающих станций для возбуждения или торможения. Кроме ганглиозной клетки, на рис. 1.4 показаны другие виды нейронов. Термины для описания структуры нейрона, в частности дендритов, несколько спорны, но, тем не менее, они удобны и широко применяются.
Не все нейроны соответствуют простому строению клетки, показанному на рис. 1.4. У некоторых нейронов нет аксонов; у других есть аксоны, на которых образуется связь. Есть клетки, чьи дендриты могут проводить импульсы и образовывать связи с клетками-- мишенями. Если ганглиозная клетка соответствует схеме стандартного нейрона с дендритами, телом и аксоном, то другие клетки не соответствуют этому стандарту. Например, у фоторецепторов (рис. 1.2С) нет очевидных дендритов. Активность фоторецепторов не вызывается другими нейронами, но активируется внешними стимулами, освещением. Другим исключением в сетчатке является отсутствие, аксонов у фоторецепторов.
Методы идентификации нейронов и прослеживание их связей
Хотя техника Гольджи все еще широко используется, многие новые подходы облегчили функциональную идентификацию нейронов и синаптических связей. Молекулы, которые окрашивают нейрон полностью, могут быть инъецированы через микропипетку, которая одновременно регистрирует электрический сигнал. Флуоресцентные маркеры, такие как люцифер желтый, позволяют увидеть самые тонкие отростки в живой клетке. Внутриклеточно могут быть введены такие маркеры, как фермент пероксидазы хрена (ПХ) или биоцитин; после фиксации они образуют плотный продукт или ярко светятся в флуоресцентном свете. Нейроны можно окрасить пероксидазой хрена и при экстраклеточной аппликации; фермент захватывается и транспортируется в тело клетки. Флуоресцентные карбоциановые красители при соприкосновении с мембраной нейрона растворяются и диффундируют по всей поверхности клетки. Эти приемы
Глава 1. Передача информации и структурная организация мозга |
19 |
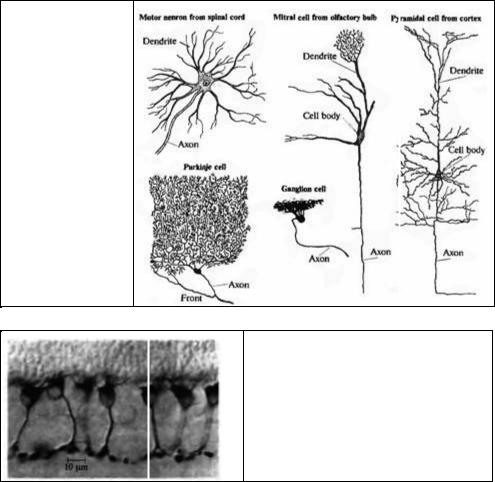
Рис. 1.4. Формы и размеры нейронов.
Ид. 1.4. Shapes and Sizes of Neurons. Neurons have branches (the dendrites) on which other neurons form synapses, and axons that in turn make connections with other neurons. The motor neuron, drawn by Deiters in 1869, was dissected from a mammalian spinal cord. The other cells, stained by the Golgi method, were drawn by Ramon y CajaL The pyramidal cell is from the cortex of a mouse, the mitral cell from the olfactory bulb (a relay station in the pathway concerned with smell) of a rat the Purkinje cell from human cerebellum, and the ganglion cell from mammalian retina (animal not specified). (After Ramon y CajaL 1995.)
Рис. 1.5. Группа биполярных клеток, окрашенных антителом на фермент фосфокиназа С. Только содержащие фермент клетки окрасились.
Fig. 1.5. Population of Bipolar Cells Stained by an Antibody against the enzyme phosphokinase C. Only bipolar cells that contain the enzyme are stained. Above are photoreceptors; below are ganglion cells. (Photograph kindly provided by H. M. Young and D. I. Vaney, University of Queensland.)
очень важны для прослеживания прохождения аксонов из одной части нервной системы в другую.
Для описания специфических нейронов, дендритов и синапсов путем избирательного маркирования внутриклеточных или мембранных компонентов используют антитела. На рис. 1.5 показана группа специфических биполярных клеток, маркированных антителом к ферменту фосфокиназа С. Антитела успешно применяются для прослеживания миграции и дифференциации нервных клеток в онтогенезе. Дополнительным подходом для описания нейронов является гибридизация in situ: специфически меченые зонды маркируют мРНК нейрона, которая кодирует синтез канала, рецептора, передатчика или структурного элемента.
Ненервные элементы мозга
Отчетливо видна клетка, помеченная буквой M на рис. 1.2В, представляющая собой ненервную клетку, находящуюся в сетчатке. Такие клетки известны как глиальные клетки. В отличие от нейронов, у них нет аксонов или дендритов и они не связаны напрямую с нервными клетками. Глиальных клеток очень много в нервной системе. Они выполняют много разных функций, связанных с передачей сигнала. Например, аксоны ганглиозных клеток сетчатки, составляющие оптический нерв, проводят импульсы очень быстро, потому что они окружены изолирующей липидной оболочкой, называемой миэлин. Миэлин формируется глиальными клетками, которые оборачиваются вокруг аксонов при онтогенетическом развитии. Глиальные клетки сетчатки известны как мюллеровские клетки.
20 |
Раздел I. Введение |
Группировка клеток в соответствии с функцией
Замечательным свойством сетчатки является расположение клеток в соответствии с функцией (см. рис. 1.2). Клеточные тела фоторецепторов, горизонтальных, биполярных, амакриновых и ганглиозных клеток расположены отчетливыми слоями. Подобная слоистость наблюдается повсеместно в мозге. Например, структура, в которой волокна оптического нерва заканчиваются (латеральное коленчатое тело), состоит из 6 слоев клеток, которые легко различить даже невооруженным глазом. Во многих областях нервной системы клетки со сходными функциями сгруппированы в отчетливые шарообразные структуры, известные как ядра (не путайте с ядром клетки) или ганглии (не путайте с ганглиозными клетками сетчатки).
Подтипы клеток и функция
Упрошенное представление структуры сетчатки на рис. 1.2 не отражает некоторых свойств сетчатки. Существует несколько отчетливых типов ганглиозных, горизонтальных, биполярных и амакриновых клеток, каждый из которых обладает характерной морфологией, специфичностью медиатора и физиологическими свойствами. Например, фоторецепторы разделяются на два легко различимых класса — палочки и колбочки, — которые выполняют различные функции. Удлиненные палочки исключительно чувствительны к малейшим изменениям в освещении. Когда вы читаете эту страницу, рассеянный свет слишком ярок для палочек, которые функционируют только в слабом свете после длительного периода в темноте. Колбочки отвечают на зрительные стимулы в ярком свете. Более того, колбочки далее подразделяются на подтипы фоторецепторов, чувствительные к красному, зеленому или синему цвету. Амакриновые клетки являются ярким примером клеточного разнообразия: более 20 типов может быть выделено по структурным и физиологическим критериям.
Таким образом, сетчатка иллюстрирует глубочайшие проблемы современной нейробиологии. Неизвестно, для чего нужно столько типов амакриновых клеток и какие разные функции выполняет каждый из этих типов клеток. Отрезвляет сознание того, что функция подавляющего большинства нервных клеток центральной, периферической и висцеральной нервной системы неизвестна. В то же время это неведение подсказывает, что многие основные принципы роботы мозга еще не поняты.
Конвергенция и дивергенция связей
Стрелками на рис. 1.2А показано направление передачи сигнала от рецепторов к ганглиозным клеткам. В реальности картина существенно более сложная. Например, наблюдается сильное уменьшение количества вовлеченных клеток на пути от рецепторов к ганглиозным клеткам. Выходы более чем 100 миллионов рецепторов конвергируют на 1 миллионе ганглиозных клеток, аксоны которых составляют оптический нерв. Таким образом, многие (но не все) ганглиозные клетки получают входы от большого количества фоторецепторов (конвергенция) через вставочные клетки. В свою очередь, одна ганглиозная клетка интенсивно ветвится и оканчивается на многих клетках-мишенях (дивергенция, см. рис. 1.13).
Кроме того, в отличие от упрошенной схемы на рис. 1.2А, стрелки должны показывать в стороны для обозначения взаимодействия между клетками в одном слое (латеральные связи) и даже в противоположные стороны — например, назад от горизонтальных клеток к фоторецепторам (возвратные связи). Такие конвергентные, дивергентные, латеральные и возвратные влияния являются постоянными свойствами большинства нервных путей по всей нервной системе. Таким образом, простая пошаговая обработка сигнала затруднена параллельными и обратными взаимодействиями.
§ 2. Сигналы нервных клеток
Для анализа событий во внешнем мире или внутри нашего тела, для передачи информации от клетки к клетке нейроны используют электрические и химические сигналы. Расстояние передачи сигнала может быть большим: от кончиков пальцев на ногах до спинного мозга. Различные сигналы прекрасно представлены все в той же сетчатке. В то время, когда Рамон-и- Кахаль рисовал стрелки на рис. 1.2А, почти не было информации об этих сигналах, что делает его достижения еще более примечательными.
Глава 1. Передача информации и структурная организация мозга |
21 |
Ступени переработки информации можно последовательно проследить: свет падает на фоторецепторы и генерирует электрические сигналы, которые воздействуют на биполярные клетки. От биполярных клеток сигналы передаются к ганглиозным клеткам и от них к высшим центрам мозга, которые и осуществляют восприятие внешнего мира. В следующих разделах рассматриваются свойства сигналов и пути переработки информации.
Классы электрических сигналов
Электрические сигналы нервных клеток могут быть разделены на два основных класса. Вопервых, это локальные градуальные потенциалы (см. рис. 1.8), которые вызываются такими внешними стимулами, как свет, падающий на фоторецепторы глаза, звуковая волна, деформирующая волосковые клетки уха, или прикосновение, механически смещающее отросток сенсорной клетки в коже. Сходны по характеристикам, но существенно отличаются по происхождению сигналы, генерируемые в синапсах — соединениях между клетками, которые мы обсудим позже. Все эти сигналы градуальны и привязаны к месту возникновения, а их распространение зависит от пассивных характеристик нервных клеток.
Потенциалы действия составляют вторую основную категорию (см. рис. 1.9). Потенциалы действия вызываются локальными градуальными потенциалами. В отличие от локальных потенциалов, они быстро распространяются на большие расстояния — например, от глаза до высших центров по волокнам ганглиозных клеток, составляющих оптический нерв, или от моторных клеток в спинном мозге к мышцам ноги. Второе отличие потенциалов действия состоит в том, что они фиксированы по амплитуде и длительности, как точки в азбуке Морзе. Крайне существенно понимать, что потенциалы действия, путешествующие по волокнам оптического нерва не являются эпифеноменами, присутствующими лишь в наших представлениях о работе мозга. Они являются единственной формой сигнализации, которая снабжает мозг информацией о внешнем мире.
Передача сигнала от сетчатки может быть представлена следующей упрощенной схемой:
Свет
↓
Локальный градуальный сигнал в фоторецепторе
↓
Локальный градуальный сигнал в биполярной клетке
↓
Локальный градуальный сигнал в ганглиозной клетке
↓
Потенциал действия в ганглиозной клетке
↓
Проведение сигнала к высшим центрам.
Универсальность электрических сигналов
Важным свойством электрических сигналов является то, что они фактически идентичны во всех нервных клетках организма независимо от того, запускают ли они движение, передают ли информацию о цветах, формах или болезненных стимулах, или соединяют различные области мозга. Вторым важным свойством сигналов является то, что они настолько одинаковы у разных животных, что даже умудренный опытом исследователь не способен точно отличить запись потенциала действия от нервного волокна кита, мыши, обезьяны или профессора. В этом смысле потенциалы действия могут считаться стереотипными единицами. Они являются универсальным эталоном для обмена информацией во всех исследованных нервных системах. В мозге не типы сигналов, а огромное количество клеток (от 1010 до 1012 нейронов) и разнообразие связей обеспечивают сложность выполняемых задач.
Эта идея была высказана в 1868 году немецким физиком и биологом Германом фон Гельмгольцем. Беря за основу гипотетические принципы, задолго до обнаружения известных сейчас фактов, он писал2):
Нервные волокна часто сравнивают с телеграфными проводами, пересекающими местность, и это сравнение хорошо приспособлено для иллюстрации удивительных и важных особенностей их образа действия. В телеграфной сети везде мы обнаруживаем те же медные или стальные провода, несущие только один вид движения, поток электричества, но вызывающие самые разные результаты на разных станциях в соответствии с дополнительной аппаратурой, с которой провода соединены. На одной станции эффект состоит в звонке колокольчика, на другой сигнал просто передается дальше, на третьей вступает в работу записывающий аппарат. ...Говоря коротко, каждое из... различных действий, вызываемых

22 |
Раздел I. Введение |
Рис. 1.6. Техника регистрации электрической активности. (А) Кончик тонкого металлического электрода расположен близко к нервной клетке в коре. (В) Внутриклеточная регистрация производится заполненной проводящей жидкостью стеклянной микропипеткой, введенной в клетку. (С) Внутриклеточное отведение также делается пэтч-пипеткой, которая прилипает к клеточной мембране.
Fig. 1.6. Electrical Recording Techniques. (A) The tip of a fine wire electrode is located close to a nerve cell in the cortex. (The wire above the tip is insulated.) Extracellular recording allows one to record from a single cell or from a group of cells. (B) Intracellular recordings are made with a fluid-filled glass capillary that has a tip of less than 1 /urn in diameter, which is inserted into a neuron across the cell membrane. At rest there is a potential difference of about 70 mV, the inside negative with respect to the outside. This difference is known as the resting potential. (C) Intracellular recordings are also made with patch electrodes. A patch electrode has a larger tip than that of an intracellular microelectrode; the tip makes an extremely tight seal with the cell membrane. If the seal is intact the currents that flow as a single ion channel in the membrane opens or closes can be recorded. Alternatively, as shown here, the cell membrane can be ruptured to allow the diffusion of molecules between the pipette and the intracellular fluid of the cell (whole-cell patch clamp).
электричеством, может быть вызвано и передана проводом в любую необходимую точку. При этом β проводе происходит один и тот же процесс, приводящий к самым разным последствиям. ...Та разница, которую мы видим при возбуждении различных нервов, заключается только в рознице самих органов, к которым присоединен нерв и которым передается состояние возбуждения.
На самом деле, как будет показано в главе 6, небольшая разница в амплитуде и длительности очевидна в потенциалах действия разных нейронов. Утверждение, что все потенциалы действия одинаковы, равносильно утверждению, что все дубы одинаковы.
Техника записи сигналов от нейронов с помощью электродов
Для решения некоторых задач существенно регистрировать активность одного нейрона или даже одного ионного канала, тогда как для других задач необходима суммарная активность многих нейронов. Ниже коротко суммируются основные приемы для записи активности нейронов, используемые для обсуждения в следующих главах.
Впервые запись потенциалов действия от нерва была сделана от периферических нервов экстра клеточным и электродами. Пропускание тока между парой серебряных проводников вызывало потенциал действия, тогда как вторая пара таких же электродов на некотором расстоянии регистрировала ответ. В центральной нервной системе регистрация от нейрона или группы нейронов производится экстраклеточным электродом, который состоит из проводника в изолирующей оболочке или из стеклянного капилляра, заполненного проводящим солевым раствором (рис. 1.6А).
С помощью внутриклеточного микроэлектрода мы можем прямо измерять разницу потенциала между наружной и внутренней средой клетки, так же как возбуждение, торможение и возникновение импульсов. Стеклянный микроэлектрод, заполненный солевым раствором и с кончиком менее 0,1 мм в диаметре, вводится в клетку с помощью микроманипулятора (рис. 1.6В). Микроэлектроды также используют для пропускания тока через мембрану или внутриклеточной инъекции молекул в цитоплазму.