

32. Системы генетических групп крови
|
Системы (локусы) |
Антигены |
Число антигенов |
|
|
Крупный рогатый скот |
|
|
А |
А, Аь А2, D, D,, D2, H, Z' |
8 |
|
В |
О?' ОаЬ И' И p'^'p^q' С?,' \S%°Y |
>40 |
|
|
У,, Y2, А', А,, Ё', Е2) Е3) Ё^ |
|
|
С |
С,, С2, С3, Е, R,, Ra, W, W,, W2 и др. |
>10 |
|
F-V |
F(Fb F2), V |
2 |
|
J |
h,h |
2 |
|
L |
L |
1 |
|
М |
Мь Ma, M', m |
4 |
|
S |
S(Sb Sa), U(U,, U2), H', U'(U',, Uz), H", S", |
10 |
|
|
U" |
|
|
Z |
Z(Z,, Z2) |
1 |
|
R'—S' |
R', S' |
2 |
|
Т |
Г |
1 |
|
N' |
N' |
1 |
|
А |
Act Ap, Aq, Ay/, Ax |
5 |
|
В |
Ba, Вь |
2 |
|
С |
|
3 |
210
211
|
Продолжение | ||
|
Системы (локусы) |
Антигены |
Число антигенов |
|
D |
Da, Db, Dc |
3 |
|
Е |
Еа, Еь, Ed, Ee, Ef, Е,, Eh, Ei, Ej, Ek, ELE™ Еш Eo, Ep, Er, Es, Et |
18 |
|
F |
Fa, Fb, Fc> Fd |
4 |
|
G |
Ga, Gb, Gc, Gd |
4 |
|
Н |
На, Нь, He, Hd, He |
5 |
|
I |
^a, 'b |
2 |
|
J |
Ja, Jb |
2 |
|
К |
•tr- T/" ■жг лг Tf Yf Tf Tf JVq, Л^), J^c, r^t Л{1 ^f( J4g> Aq |
8 |
|
L |
La, U, Lc, Ld, Le, Lf, Ц, U, U, Lj, Lk, LI, Lm |
13 |
|
М |
Ma, Mb, Mc, Md, Me, Mf, Mg, Mh, Mi, Mj, Mk |
11 |
|
N |
Na, Nb, Nc |
3 |
|
<Ч |
Oa, Ob |
2 |
|
Р |
Pa, Po |
2 |
|
Q |
Qa, Qo |
2 |
Антигены некоторых систем наследуются в определенных комбинациях — феногруппах. Например, сложная система Е у свиней включает 18 антигенов. Феногруппа Ebdg определяется присутствием антигенных факторов Еь, Ed, Eg. В этом случае аллель записывают EW8. Антигенные факторы системы В у крупного рогатого скота В, G и К могут встречаться в комбинациях В, G, BG, BGK, а аллели обозначаются Вв, BG, BBG и BBGK. В феногруппу может входить до 10 антигенов. Для упрощения записи феногруппы кодируют. Так, феногруппу BGKO2YiA'B'E'G'K'O'Y' обозначают В28.
Наследование групп крови
У всех видов животных большинство аллелей генетических систем групп крови наследуется по типу кодоминирования, т. е. в гетерозиготе фенотипически проявляются оба гена. Весьма редко встречаются рецессивные аллели, подобные аллелю О системы АВО у человека. В связи с этим возможен анализ частоты аллелей разных локусов в популяциях во времени и в пространстве, что является главным инструментом для описания их генетической структуры и позволяет приблизиться к пониманию эволюционного процесса.
Все известные системы групп крови у сельскохозяйственных животных локализованы в аутосомах. В сложных системах (у скота В- и С-системы) антигенные факторы контролируются несколькими тесно сцепленными сублокусами. С-система состо-
212
ит из двух серий аллельных (или почти близко к аллельным) генетических детерминант Сь С2, С'{ ,С2 и Хь Х2) С, Fi0. Анализ рекомбинаций между концевыми антигенами С-системы показал, что длина участка ДНК этой системы составляет 0,3 сМ (сантиморгана), тогда как В-системы — 0,7 сМ.
Можно выделить три основных правила наследования групп крови: 1) каждая особь наследует по одному из двух аллелей от отца и от матери в каждой системе групп крови; 2) особь с антигенами, не обнаруженными хотя бы у одного из родителей S^^SST быть потомком данной родительской пары (например, Р $ Fp/F х d'FF/V * FiN/N); 3) гомозиготная особь по одному антигену, например F/F, не может быть потомком гомозиготной особи с противоположным антигеном, например V/V.
Получение реагентов для определения групп крови. Иммунологическая специфичность белковых антигенов определяется: 1) последовательностью аминокислот полипептидной цепи (иногда изменение даже нескольких аминокислот ведет к изменению антигена); 2) концевыми аминокислотами цепи; 3) вторичной структурой белковой молекулы; 4) наиболее активными поверхностно расположенными участками полипептидной цепи — антигенными детерминантами (один антиген может иметь несколько детерминантных групп).
Антигены выявляются при помощи реакции антиген — антитело. Основой для определения взаимодействия антиген — антитело служит у крупного рогатого скота и овец реакция гемолиза (разрушение стромы эритроцитов с выделением из них гемоглобина), у свиней — полная и неполная агглютинация (склеивание эритроцитов) и реакция гемолиза.
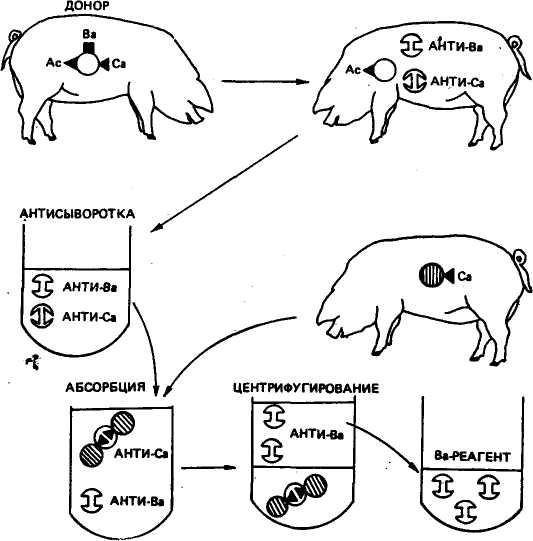
Схема получения моноспецифической сыворотки В приведена на рисунке 42. Кровь от животного-донора, имеющего антигены Ас, Ва и Са, вводят реципиенту с антигеном Ас, но не имеющему антигенов Ва и Са. У реципиента вырабатываются антитела к антигенным факторам Ва и Са. Антитела против антигена Ас не образуются, так как у реципиента есть этот фактор. В сырой сыворотке абсорбируют ненужные антитела, в данном случае анти-Са, эритроцитами третьего животного, имеющего антиген Са. Потом из сыворотки путем центрифугирования удаляют эритроциты с абсорбированными на них антителами Са. Полученную моноспецифическую сыворотку можно использовать для выявления антигена Ва в эритроцитах других животных.
Системы групп крови. В настоящее время у крупного рогатого скота открыто 12 систем групп крови, у свиней — 17, у овец — 16, у лошадей — 9, у птиц — 14. Из всех этих систем наиболее сложной является В-система у крупного рогатого скота, включающая более 40 антигенов, которые в различных комбинациях образуют более 500 аллелей. Если в системе имеется более трех
213
РЕЦИПИЕНТ
ЭРИТРОЦИТЫ С АНТИГЕННЫМ ФАКТОРОМ Ас
ЭРИТРОЦИТЫ С АНТИГЕННЫМ ФАКТОРОМ Ва
ЭРИТРОЦИТЫ С АНТИГЕННЫМ ФАКТОРОМ Са 5£ АНТИТЕЛА ПРОТИВ АНТИГЕНА В В СЫВОРОТКЕ 5J АНТИТЕЛА ПРОТИВ АНТИГЕНА С В СЫВОРОТКЕ
Рис. 42. Схема получения моноспецифической сыворотки путем иммунизации
аллелей, то такие системы называют полиаллельными. К ним кроме системы В относят системы С, S, А, у свиней — Е, L, М, у овец — В, А, С.
J-система крупного рогатого скота имеет иммуногенетическое сходство с антигеном А человека, свиней и антигеном R овец, S-система гомологична М-системе овец. Система Р групп крови у лошади аналогична АВО-системе человека. У крупного рогатого скота установлена связь J-системы с локусоМ| гемоглобина (НЬ) и Р-лактоглобулина (pL)
ЗНАЧЕНИЕ ГРУПП КРОВИ ДЛЯ ПРАКТИКИ
Контроль достоверности происхождения животных. Одна из главных областей практического применения групп крови — контроль происхождения животных. Такое их использование вызвано тем, что в некоторых стадах встречается 20 % и более ошибок в происхождении животных. Это может быть следствием не только недостатков в работе техников по искусственному осеменению, потери номеров, неправильного их чтения, но и результатом повторных осеменений животных спермой разных производителей (в повторную охоту приходит до 50 % коров, а продолжительность стельности в норме изменяется от 270 до 292 дней) и других случаев.
Контроль происхождения необходим и при испытании свиноматок по качеству потомства, осемененных смешанной спермой хряков (В. Н. Тихонов, 1967), для установления моно- и дизи-готности двоен, при получении животных методом трансплантации эмбрионов и т. д.
Контроль достоверности происхождения животных возможен благодаря: 1) кодоминантному наследованию антигенных факторов; 2) их неизменности в течение онтогенеза; 3) огромному числу комбинаций групп крови, которые в пределах вида практически не бывают одинаковыми у двух особей, за исключением монозиготных близнецов.
В таблице 33 приведен пример уточнения отцовства в случае, когда корова в первый раз и повторно была осеменена спермой разных быков. По системе А невозможно уточнить происхождение потомка, так как аллель DH есть у обоих быков. В системе В теленок получил один аллель ВО2А от матери (такого аллеля нет у предполагаемых отцов), а второй АВ — от быка № 2 (этого аллеля нет у первого производителя). Поэтому уже можно сделать заключение, что отцом теленка является бык № 2 (исходя из второго правила). Это заключение подтверждается и наличием у потомства аллеля W в системе С. Точно так же по системе F—V можно сделать заключение, что первый производитель не может быть отцом, так как он гомозиготен по аллелю F/F, а потомок гомозиготен по противоположному аллелю V/V (третье правило).