

Глава 5 генетика пола
На основании анализа результатов многочисленных экспериментов с дрозофилой Т. Морган сформулировал хромосомную теорию наследственности, сущность которой заключается в следующем: 1) гены находятся в хромосомах, располагаются в них линейно на определенном расстоянии друг от друга; 2) гены, расположенные в одной хромосоме, относятся к одной группе сцепления. Число групп сцепления соответствует гаплоидному числу хромосом; 3) признаки, гены которых находятся в одной хромосоме, наследуются сцепленно; 4) в потомстве гетерозиготных родителей новые сочетания генов, расположенных в одной паре хромосом, могут возникать в результате кроссинговера в процессе мейоза. Частота кроссинговера зависит от расстояния между генами; 5) на основании линейного расположения генов в хромосоме и частоты кроссинговера как показателя расстояния между генами можно построить карты хромосом.
Контрольные вопросы. 1. В чем заключается сцепленное наследование признаков и как оно устанавливается? 2. Сколько групп сцепления у разных видов сельскохозяйственных животных? 3. Как наследуются признаки при полном и неполном сцеплении? 4. Что такое кроссинговер? Когда и как он происходит и от чего зависит его частота? 5. Как определяется частота кроссинговера? Почему она принята за единицу расстояния между генами? 6. В чем состоит биологическое значение кроссинговера? 7. Как было доказано, что гены в хромосоме расположены линейно? 8. Что такое карта хромосомы и как она составляется? 9. Что такое соматический кроссинговер? 10. В чем заключается хромосомная теория наследственности?
ДЕТЕРМИНАЦИЯ ПОЛА И МЕХАНИЗМ ЕГО НАСЛЕДОВАНИЯ
В ходе эволюции у большинства раздельнополых организмов сформировался механизм детерминации пола, обеспечивающий образование равного количества самцов и самок, что необходимо для нормального самовоспроизведения вида. Детерминация пола может происходить на разных этапах размножения. Различают три основных типа детерминации:
1) эпигамный, когда пол особи определяется в процессе онто-йн^за^Детерминация пола в данном случае значительно зависит от внешней среды; j
^УЪЫй когда пол будущего дочернего организма опре б
Ур, уу р
деляется в ходе гаметогенеза у родителей особи;
3) сингамный, при котором пол дочерней особи определяется в момент слгоШия гамет. Это наиболее распространенный тип детерминации пола, характерный для животных всех видов.
При прогамном и сингамном типах детерминации пол зависит от определенных половых хромосом.
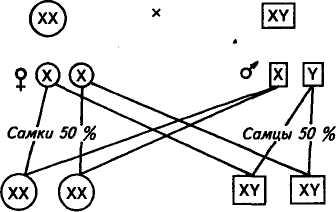
У самок млекопитающих в диплоидном наборе хромосом выделяют пару одинаковых по форме гоносом (половых хромосом), обозначаемых ХХ-хромосомами. Самцы в кариотипе содержат Х- и Y-хромосомы. Самки птиц содержат две разные (XY), а самцы одинаковые (XX) половые хромосомы. В период редукционного деления (мейоза) у самок млекопитающих образуется один тип гамет с Х-хромосомой, поэтому женский пол называют гомогаметным. У самцов образуется два типа гамет с Х- и Y-хромосомами, поэтому мужской пол называют гетерогаметным. Определение пола млекопитающих зависит от того, каким спермием будет оплодотворена яйцеклетка. Если яйцеклетка оплодотворена спермием, содержащим Х-хромосому, то происходит закладка особи женского пола, если спермий несет Y-хромосому — закладывается особь мужского пола.
Схема образования гамет в мейозе самцов и самок, определения пола при оплодотворении следующая:
63
p
Гаметы Потомство
Существует равная вероятность закладки особи мужского или женского пола, так как число сперматозоидов у самцов (яйцеклеток у птиц) с Х- и Y-хромосомами одинаково.
Самцов обычно рождается на несколько процентов больше, чем самок, но в ходе эволюции выработался механизм сохранения числового соотношения полов, поэтому к возрасту половой и физиологической зрелости это соотношение выравнивается вследствие более высокой смертности самцов.