

Molecular and Cellular Signaling - Martin Beckerman
.pdf318 13. Cell Fate and Polarity
FIGURE 13.8. Hedgehog receptors Patched and Smoothened: (a) Patched receptor.
(b) Smoothened receptor in a closed conformation. (c) Smoothened receptor in an open configuration.
events. For example, the Hh pathway is responsible for body, wing, eye, genitals, and leg patterning in Drosophila while the Shh pathway regulates eye, portions of the brain, hair, lung, gut, bladder, and urethra patterning in mammals.
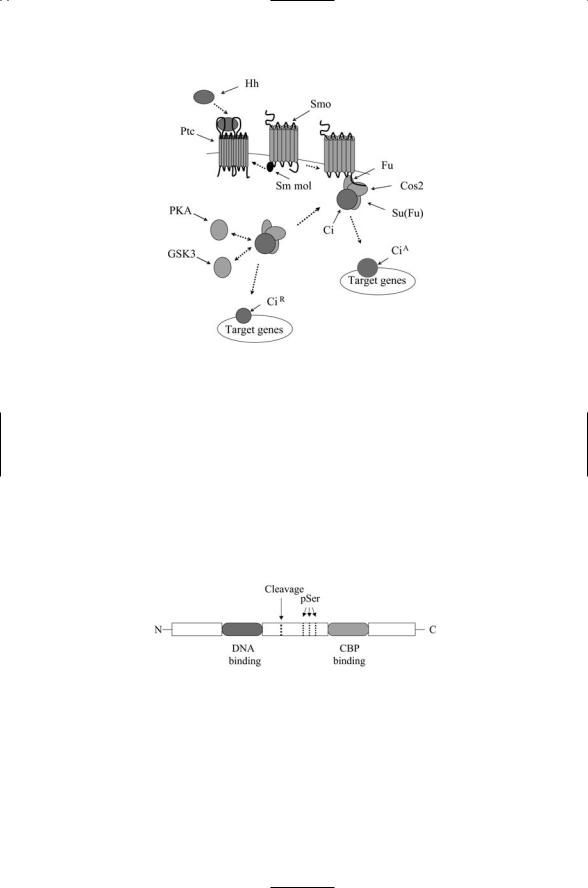
The Patched receptor is a 12-pass transmembrane protein (Figure 13.8). It functions together with a partner receptor called Smoothened. The Smoothened protein is a 7-pass transmembrane receptor similar to the Wnt receptor Frizzled. Patched is responsible for ligand binding and Smoothened is responsible for transducing the signal across the plasma membrane. Two different conformations of the Smoothened receptor are depicted in Figure 13.8. In the first, (b), the cytoplasmic tail cannot bind to the downstream signaling partners, but, in the second, (c), binding sites along the cytoplasmic tail become available. In the absence of ligand binding, Patched acts catalytically to maintain Smoothened in its closed conformation, but ceases to do so when bound to Hedgehog.
The cytoplasmic tail (CT) of Smoothened in its open conformation interacts with two cytoplasmic proteins—Costal-2 (Cos2) and Fused (Fu). The first of these, Cos2, is a kinesin-like protein that binds to microtubules while the other, Fu, is a serine/threonine kinase. The Cos2 protein functions as a scaffold for assembly of several signaling elements. These include, besides Smoothened, CT and Fused, Suppressor of Fused (Su(Fu)) and a down- stream-acting transcription factor called Cubitus interruptus (Ci). Two additional serine/threonine kinases—PKA and GSK3—complete the specification of this Hedgehog signaling node (Figure 13.9).
13.9 Gli Receives Hh Signals
The primary recipients of Hh signals are Gli transcription factors. Glis (so named because of their involvement in malignant gliomas) are zinc finger, DNA-binding proteins. They have five zinc fingers, three of which grip the DNA molecule, a CBP binding domain, and numerous phosphorylation sites (Figure 13.10). The best studied of these proteins is the Drosophila Cubitus interruptus protein. As just described this protein forms a complex
13.9 Gli Receives Hh Signals |
319 |
FIGURE 13.9. Signaling through the Hedgehog pathway: In the absence of ligand binding, Patched (Ptc) represses Smoothened (Smo), depicted in the figure as acting through a small molecular (Sm mol) intermediary. In response to ligand binding, the repression is relieved and Smo undergoes a conformational change from a closed configuration to an open one that can bind Cos2 and Fu. This binding action prevents phosphorylation of Ci by PKA and GSK3. The complex dissociates and the full length 155 kDa CiA translocates to the nucleus where it promotes transcription. In the absence of ligand binding, the Ci protein is cleaved following phosphorylation, and the 75 kDa CiR protein translocates to the nucleus where it represses transcription.
FIGURE 13.10. Organization of the Drosophila Cubitus interruptus (Ci) protein: Five tandem zinc fingers situated in the N-terminal half of the protein are responsible for binding DNA. A CBP binding domain located in the C-terminal portion of Ci binds a CBP cofactor. Cleavage of the protein produces the CiR protein containing the N-terminal portion and acting as a transcriptional repressor. Three of the many S/T phosphorylation sites (pSer) are shown. The sites indicated are the priming sites phosphorylated by protein kinase A (PKA). Sites located both N-terminal and C-terminal to these sites are phosphorylated by GSK3.

320 13. Cell Fate and Polarity
with Cos2, Fu, and Su(Fu) that is bound to microtubules. In the absence of Hh signaling, another member of the signaling pathway, protein kinase A (PKA) phosphorylates Ci. This phosphorylation event tags Ci for proteolysis. The result of this process is the formation of a 75-kDa CiR protein (Figure 13.9), which translocates to the nucleus where it functions as a repressor of the transcription of several genes. When Hh is present, phosphorylation of Ci by PKA is blocked, the complex dissociates, and the fulllength 155-kDa protein is left intact and free to move into the nucleus. This intact CiA protein functions as an activator of transcription of a number of genes including patched, dpp, and wg.
13.10Stages of Embryonic Development Use Morphogens
The fertilized egg of multicellular animals goes through a sequence of developmental stages. The egg first goes through a cleavage stage where it divides mitotically several times to form a ball of smaller cells, or blastomeres. In the next phase, the cells move to the outside forming an epithelial sheet that encloses a fluid-filled chamber. In the third stage, gastrulation, the single-layered blastula develops into a gastrula consisting of three layers of cells—the endoderm, mesoderm, and ectoderm, roughly corresponding to gut, connective tissue and muscles, and epidermis (respectively) of the adult organism.
In more detail, the lungs, components of the digestive system such as stomach and liver, and associated glands and structures, develop from the endoderm. In the developing mesoderm, left and right sides of the body are delineated by a notocord that defines the central axis of the body. Sections of mesodermal cells progressively bud off on both sides of the notocord to form somites, which then develop further into the individual vertebra and muscle groups. The vascular system, including the heart, bone, and cartilage, develop from the mesoderm, while the nervous system and associated sensory organs develop from the ectoderm in a developmental stage called neurulation.
The term morphogen was introduced at the beginning of the chapter. Morphogens are signaling proteins that are expressed either on cell surfaces or secreted into the extracellular spaces in the form of concentration gradients. These gradients are subsequently “read” by cells to determine their developmental fate. Cells adopt different cell fates according to their position in the gradient relative to the signal source.
Cells in organizing centers, at boundaries between different layers or regions, and within regions, in the embryo secrete morphogens during development. Organizing centers are localized groupings of one or more kinds of cells that secrete morphogens in order to impart patterns of cell fates to fields of progenitor cells. The morphogens are secreted not only at specific

13.11 Gene Family Hierarchy of Cell Fate Determinants in Drosophila |
321 |
locations but also at specific times during embryonic development. These morphogens and associated signaling proteins are expressed sequentially; that is, through a hierarchical pattern of gene expression with each family of gene products preparing the way for the next family of gene products.
13.11Gene Family Hierarchy of Cell Fate Determinants in Drosophila
In Drosophila, five sets of gene products have major roles in determining cell fate. The gene families form a hierarchy with one set of gene products preparing the way for the next set of genes and their protein products. Each set contributes to the emergence of the body plan, producing a succession of progressively finer partitions of the embryonic body into segments and compartments that eventually become adult body parts such as head, thorax, abdomen, wings, and legs. Although the details differ from phylum to phylum these families are highly conserved among multicellular organisms up to an including vertebrates. These families, their functions, and members are presented in Table 13.3.
The earliest set of gene products is the maternal effect genes. As their name indicates, they are supplied maternally. Some of these gene products function as morphogens, while others assist in morphogen localization. The morphogen gradients are generated internally within the single cell, the egg, rather than externally by many cells, and contribute to the establishment of cell polarity and asymmetric cell division. Maternal effect genes prepare the cell for expression of the gap genes. These are distributed in broad bands
TABLE 13.3. Hierarchy of Drosophila patterning genes.
Gene family |
Function |
Members |
Maternal effect |
Establish anterior-posterior and |
A/P axis: Bicoid, caudal, |
|
dorsal-ventral body axes; |
hunchback, nanos, oskar, |
|
regulate gap and pair rule |
stauffen; D/V axis: cactus, |
|
gene expression pattern |
dorsal, pelle, spätzle, toll, |
|
|
tube |
Gap |
Partition body into three broad |
Zygotic caudal, giant, zygotic |
|
regions; regulate pair rule |
hunchback, huckebein, |
|
gene expression |
knirps, krüppel, tailless |
Pair rule |
Partition body into bands; |
Even-skipped, fushi tarazu, hairy, |
|
regulate segmentation gene |
odd-paired, odd-skipped, |
|
expression |
paired, runt, sloppy paired |
Segment |
Establish anterior-posterior |
Armadillo, cubitus interruptus, |
polarity |
axis within each |
engrailed, frizzled, fused, |
|
compartment |
hedgehog, patched, wingless |
Hox cluster |
Establish body part identity |
Abd-A, Abd-B, Antp, Dfd, Lab, |
|
|
Pb, Scr, Ubx |
|
|
|

322 13. Cell Fate and Polarity
to anterior, middle, and posterior regions of the embryo. If a gap gene is missing the corresponding portion of the body does not develop, producing a gap, and hence the name. These genes prepare the way for the expression of pair rule genes. The pair rule genes further partition the broad segments produced by the gap genes into seven embryonic segments, or parasegments. If one of these genes is missing every other segment in the developing larva is absent, and hence the name “pair rule.” The next set of genes, segment polarity genes, specify the polarity within each parasegement; that is, they define the anterior-posterior axes of each of the parasegments. Finally, the Hox genes help delineate the various adult body parts.
13.12 Egg Development in D. Melanogaster
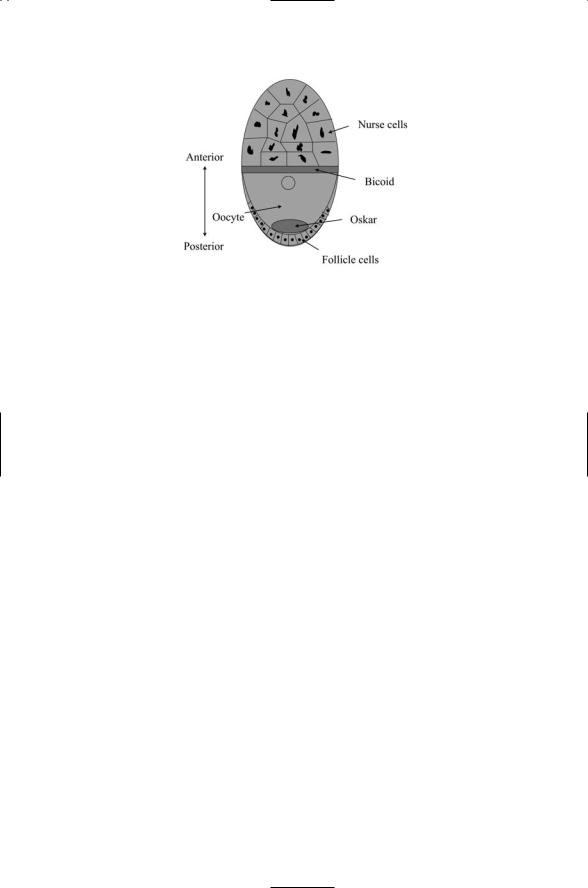
The establishment of asymmetry and polarity in D. melanogaster begins prior to fertilization, during the development of the egg. In the egg development (oogenesis) stage, an initial germ cell divides four times, producing a single egg cell and 15 nurse cells. The nurse cells are connected to the egg by cytoplasmic bridges that permit mRNAs to flow into the egg cell from the nurse cells. Somatic follicle cells surround and nurture the resulting assemblage. Whereas the point of entry of the sperm into the egg determines the polarity of the C. elegans zygote, the orientation of the Drosophila oocyte is determined by signals exchanged between it and the most posterior follicle cell. The Gurken protein plays a key role in this signaling pathway, which involves two-way signaling between the egg cell and its surrounding cells in which directional information is encoded through a polarized microtubule cytoskeleton.
Once an initial orientation is specified through communication with the neighboring cells, distributions of mRNAs and proteins are set up in regions of the egg and zygote delineated by anterior/posterior and dorsal/ventral axes. Some mRNAs and proteins are localized either to the anterior or posterior regions of the cell, while others are localized in a dorsal/ventral manner. The proteins being localized belong to the signal transduction pathways and include most importantly a considerable number of transcription factors. When the cells divide, the programs of gene expression in the daughters differ from one another because of the asymmetric distributions of transcription factors, and the daughters have different cell fates.
Two of the proteins involved in delineating the anterior and posterior regions are Bicoid and Oskar. Bicoid localizes to the anterior pole and
Oskar protein collects near the posterior pole (Figure 13.11). These proteins, along with several others, guide the formation of graded distributions of mRNA for Hunchback and Caudal in the anterior and posterior regions, respectively. These last-named proteins are intracellular morphogens and their graded distributions help determine cell fate. Genes expressed in the
13.13 Gap Genes Help Partition the Body into Bands |
323 |
FIGURE 13.11. Schematic depiction of a Drosophila oocyte: Shown in the figure are the polynuclear nurse cells, oocyte, and posterior array of follicular cells. Bicoid mRNAs are laid down in a strip in the anterior end and diffuse towards the posterior pole to form a concentration gradient. Oskar mRNAs and proteins are laid down at the posterior pole and diffuse out to form a countergradient.
oocyte, acting as they do in the absence of paternal gene products, are referred to as “maternal effect genes.” The bicoid and oskar genes are maternal effect genes acting to establish cell polarity. Bicoid and Nanos, another maternal effect gene product, act as morphogens. Bicoid is laid down to form a gradient peaked at the anterior pole, while Nanos is laid down opposite way to Bicoid, having its highest concentration in the posterior pole.
Maternal effect gene products belonging to the Toll pathway are involved in delineating the dorsal/ventral distributions. Recall that the Toll pathway is an evolutionary ancient pathway that mediates innate immune responses.
Drosophila counterparts to the mammalian Toll gene products are involved in establishing the early dorsal/ventral patterning. Spätzle is the Drosophila Toll ligand; Cactus is the Drosophila counterpart to the IkB inhibitor, and Dorsal the counterpart to the NF-kB transcription factor. After fertilization occurs, the Toll signal cascade is initiated resulting in activation of Dorsal in a graded fashion due to ventral restriction of Spatzle signaling. Tube and Pelle are intermediaries in the Toll signaling pathway.
13.13 Gap Genes Help Partition the Body into Bands
Gap genes are expressed in broad regions of the developing Drosophila embryo. Maternal effect gene products initiate the expression of these genes. Working in concert the Hunchback, Knirps, Giant, and Krüppel genes products subdivide the embryo into regions along the anterior posterior axis. As shown in Figure 13.12, Krüppel is expressed in a wide band in the

324 13. Cell Fate and Polarity
FIGURE 13.12. Patterns of distributions of developmental genes in the Drosophila embryo: (a) Distribution of gap gene products Krüppel, Giant, and Knirps in broad bands. (b) Distribution of pair rule gene product Even-skipped (Eve) in the Drosophila embryo. Shown is the 7-striped zebra pattern of eve gene expression. Stripes are numbered from the anterior end to the posterior end.
center of the embryo. Knirps is expressed the in a broad posterior region adjacent to the central Krüppel region, and Giant genes are expressed on the two outer sides of the centrally situated Krüppel and Knirps regions. A second, small Knirps region is situated near the anterior pole (not shown in the figure).
Tailless and huckebein are terminal gap genes. They are expressed at both the anterior pole and posterior pole to complete the A/P partitioning of the embryo. Hunchback inhibits expression of posterior gap genes knirps and giant in the anterior part of the embryo. If hunchback is not expressed the head will be missing; if Knirps is missing the abdomen will be missing, and if Krüppel is missing the thorax and abdomen will not appear. Thus, the proteins encoded by these genes are cell fate determinants that divide the growing embryo into head, abdomen, and terminal bands.
13.14Pair-Rule Genes Partition the Body into Segments
Like the gap genes, the pair rule genes are transcription factors. Three of them—even-skipped, hairy and runt—are regarded as primary pair rule genes. The other pair rule genes are referred to as secondary pair rule genes; they are expressed later and are governed by the expression patterns of the primary pair rule genes. The pair rule genes specify segment boundaries; they are laid down in zebra-like patterns giving rise to the 7-segment striped

13.15 Segment Polarity Genes Guide Parasegment Development |
325 |
body of the developing larva. As is the case for the other primary pair rule genes, the regulatory region for even-skipped (eve) contains a set of special binding sites, called enhancer elements, for transcription factors, one for each stripe. These enhancers correspond in a unique way to the gradient information in the region of the stripes.
Gap and maternal effect genes both regulate the pair rule gene expression patterns. The gene regulatory region (enhancer) for eve in stripe 2 illustrates the general theme. Bicoid and Hunchback bind to sites in the enhancer and function as activators of gene expression. Giant and Krüppel also bind to sites in the enhancer and operate as repressors of gene expression. Since Giant is expressed at the anterior end and Krüppel in the middle, stripe 2 is inhibited in those regions but is encouraged by Bicoid and Hunchback in a stripe in the anterior region located in between the two repressed regions. Similarly, enhancers of the other stripes reflect the gradients of the corresponding sets of maternal effect and gap genes in the regions where the other stripes are to be sited. The net result is the emergence of seven stripes of expression for eve, and similarly, seven stripes for hairy and seven stripes for runt. The gene products appear in regions that partially overlap one another and so they operate in combinatorial manner to delineate the parasegments.
13.15Segment Polarity Genes Guide Parasegment Development
The development of the Drosophila larva continues with the expression of segment polarity genes. The expression of gap and pair rule genes occurs during the precellular blastomere stage of embryo development. The blastomere stage ends with cellularization where movements of the membrane about the nuclei take place, separating the nuclei and forming distinct cells.
The segment polarity genes are expressed at the start of gastrulation. Whereas the gap and pair rule gene products function as transcription factors, the segment polarity gene products include both transcription factors and signaling proteins that mediate communication between the newly formed cells and coordinate their programs of gene expression.
Genes that encode several receptor-ligand combinations appear in the list of segment polarity genes. Among the entries in Table 13.3 are patched and hedgehog, and frizzled and the gene encoding the Wnt ligand Wingless. Downstream signal transducers such as Cubitus interruptus are expressed; as well. As this stage begins Engrailed, a transcription factor, is expressed along with two secreted signaling proteins—Wingless and Hedgehog. Pair rule gene products such as Eve and Ftz establish patterns of Wg, En, and Hh gene expression, and cell-to-cell signaling sustains them. The result of this activity is the formation of 14 stripes of Engrailed/Hh gene expression and 14 stripes of Wingless gene expression.

326 13. Cell Fate and Polarity
The boundary between cells expressing Engrailed/Hedgehog and those expressing Wingless is the parasegment boundary, and the regions between pairs of boundaries are the parasegments. Cells residing in the anterior half of each parasegment express Wg and those situated in the posterior compartments express En/Hh. The cell fates in the two compartments differ: cells in the anterior compartment become “A” (anterior) cells while those in the posterior compartment become “P” (posterior) cells.
13.16Hox Genes Guide Patterning in Axially Symmetric Animals
Bilaterally symmetric animals such as humans express a family of highly conserved regulatory genes, called Hox genes. These genes play an important role in specifying which cells in the mesoderm become which body parts. In vertebrates, these genes guide the emergence of the axial skeleton (for example, the breastbone, ribcage, and spine), voluntary muscles, and dermis of the back from the somites mentioned earlier in the chapter, that is, from the repeated, identical segments of cells belonging to the mesoderm.
These genes have been studied extensively in the fly and other insects, the nematode, chick, and mouse. In all organisms studied, the genes appear as a cluster, one gene after the other. In vertebrates there are four clusters of genes, each cluster containing from 9 to 11 genes. In insects and the nematode there is a single gene cluster. The members of the Drosophila Hox cluster are listed in Table 13.3. Three of the Hox genes—Dfd, Lab, and Pb— delineate the head. Two gene products—Antp and Scr—specify the thorax, and the remaining three—Abd-A, Abd-B and Ubx—specify the abdomen.
In many metazoans, a small number of regulatory genes control the development of organs and morphological structures. These genes, called selector genes, have been intensively studied in Drosophila, where they control the formation of body parts and bilateral partitioning. These regulatory genes, of which Hox genes are prominent members, function as transcription factors. Selector genes do not work alone. Rather, they work together with other regulatory components in a combinatorial fashion to create specific patterns of gene expression, leading to the development and arrangement of body parts. The segment polarity gene, engrailed, discussed previously, is a good example of a selector gene. It works in concert with other segment polarity genes to determine which cells become A cells and which ones become P cells.
Hox genes are activated in a spatially sequential manner from anterior
(head) to posterior (tail) end. Cells progressively bud off from the main axis to form the somites. The differentiation of the cells within the somites to form vertebrae and muscles takes the form of a wave of activity whose leading edge moves down the A/P axis, with presomitic mesoderm in front of the wave and somites behind it. The movement of this wave must be care-

References and Further Reading |
327 |
fully timed. The timing activity keeps the movement of the wave in phase with the time needed for development of the somites.
References and Further Reading
Notch Signaling
Artavanis-Tsakonas S, Rand MD, and Lake RJ [1999]. Notch signaling: Cell fate control and signal integration in development. Science, 284: 770–776.
Lai EC [2002]. Keeping a good pathway down: Transcriptional repression of Notch pathway target genes by CSL proteins. EMBO Rep., 3: 840–845.
Milner LA, and Bigas A [1999]. Notch as a mediator of cell fate determination in hematopoiesis: Evidence and speculation. Blood, 93: 2431–2448.
Schroeter EH, Kisslinger JA, and Kopan R [1998]. Notch-1 signalling requires ligand-induced proteolytic release of intracellular domain. Nature, 393: 382–386.
Struhl G, and Adachi A [1998]. Nuclear access and action of Notch in vivo. Cell, 93: 649–660.
Presenilins and Matrix Metalloproteinases
De Strooper B [2003]. Aph-1, Pen-2, and Nicastrin with Presenilin generate an active g-secretase complex. Neuron, 38: 9–12.
De Strooper B, and Annaert W [2000]. Proteolytic processing and cell biological function of the amyloid precursor protein. J. Cell Sci., 113: 1857–1870.
Fortini ME [2001]. Notch and Presenilin: A proteolytic mechanism emerges. Curr Opin. Cell Biol., 13: 627–634.
Schlondorff J, and Blobel CP [1999]. Metalloprotease-disintegrins: Modular proteins capable of promoting cell-cell interactions and triggering signals by proteinectodomain shedding. J. Cell Sci., 112: 3603–3617.
Vu TH, and Werb Z [2000]. Matrix metalloproteinases: Effectors of development and normal physiology. Genes Dev., 14: 2123–2133.
TGF-b Signaling (Receptor Serine/Threonine Kinase Pathway)
Moustakas A, Souchelnytskyi S, and Heldin CH [2001]. Smad regulation in TGF-b signal transduction. J. Cell Sci., 114: 4359–4369.
Qin BY, et al. [2002]. Smad3 allostery links TGF-b receptor kinase activation to transcriptional control. Genes Dev., 16: 1950–1963.
Shi YG, and Massagué J [2003]. Mechanism of TGF-b signaling from cell membrane to the nucleus. Cell, 113: 685–700.
Ten Dijke P, et al. [2002]. Regulation of cell proliferation by Smad proteins. J. Cell Physiol., 191: 1–16.
Ten Dijke P, Miyazono K, and Heldin CH [2000]. Signaling inputs converge on nuclear effectors in TGF-b signaling. Trends Biochem. Sci., 25: 64–70.
Wnt Signaling Pathway
Cardigan KM, and Nusse R [1997]. Wnt signaling:A common theme in animal development. Genes Dev., 11: 3286–3305.
Dale TC [1998]. Signal transduction by the Wnt family of ligands. Biochem. J., 329: 209–223.