

Molecular and Cellular Signaling - Martin Beckerman
.pdf18
Regulation by Viruses
Viruses consist of nucleic acid cores encased in protein sheaths. The protein sheaths surrounding and protecting the nucleic acid cores are referred to as capsids. Viral capsids enclose either DNA or RNA. As might be expected, the viruses belonging to the former class are known as DNA viruses, while members of the latter group are called RNA viruses. Some RNA viruses covert their RNA to DNA, and from there pass through both a transcription and a translation stage to make new viruses. These viruses are called retroviruses. Other RNA viruses do not convert their RNA to DNA. Instead, their RNA functions as messenger RNA molecules that are used directly in a translation stage to synthesize proteins.
Many viruses use lipids and carbohydrates acquired from their hosts to make an envelope. They encase their caspsids in lipids and carbohydrates picked up from plasma membrane or from organelles such as the nucleus, ER, or Golgi apparatus. These viruses are referred to as enveloped viruses. Glycoproteins are an important component of the viral envelopes. These proteins mediate contact with and entry into the host cell through their ability to bind to receptors on the host’s plasma membrane.
Viral genomes vary greatly in size. Some, like those of the picornaviruses, are small, less than 10 kBp, and encode on the order of 10 proteins. Others, such as poxvirus genomes, are large. They range in size from 130 to 300 kBp and encode ~200 proteins, approaching those of the mycoplasmas in size of their genomes. In all cases, the viruses require assistance and resources from the host cell to replicate and propagate. For that reason their genomes encode not only genes of a structural character that encode proteins for the capsid and envelope, but also genes involved in replication and genes of a signaling and regulatory character. The viral signaling and regulatory genes encode proteins that regulate viral replication, disrupt and disable host immune/protective responses, and interact with cellular systems in order to use them for transport and replication.
In the first part of the chapter the viral cycle of entry, replication, and exit will be examined. This discussion will be followed by a general sketch of some of the ways viruses deal with the immune system. The remainder of
441

442 18. Regulation by Viruses
the chapter is devoted to detailed discussion of how signaling and regulatory proteins make possible the survival of specific viruses. The hepatitis C virus responsible for chronic hepatitis, cancer-causing viruses, human immunodeficiency virus (HIV)—the causative agent of acquired immunodeficiency syndrome (AIDS), and bacteriophages will be examined.
18.1 How Viruses Enter Their Host Cells
Viruses enter their host cells using receptors and the cellular machinery for membrane fusion and endocytosis. Viruses cannot pass directly through the plasma membranes of cells. Instead they possess proteins in their exposed surfaces that bind to receptors on host cells triggering their ingestion into the cell. Enveloped viruses, for example, contain proteins in their envelopes that distinguish the correct host cells from non-host cells and initiate the process whereby the virus gains entry to the cell interior. There are two main ways of entering the host cell. In endocytic entry, the virus enters the cell in vesicles associated with clathrin-coated pits and caveolae. In this mode of entry, the viral nucleic acid and the capsid are enclosed in a transport vesicle that is pinched off from the plasma membrane. In the nonendocytic mode of entry, the viral envelope fuses with the plasma membrane leading to release of the nucleic acid encased in its capsid into the cytoplasm.
In order to replicate, viruses must first gain entry into the cell, and then disassemble, replicate DNA, synthesize components, reassemble, and exit. Viruses that use eukaryotic hosts for their replication have special challenges. They must navigate in an environment containing a dense cytoskeleton and numerous organelles partitioned into many subcompartments. The viruses, even small ones, are too big to traverse intracellular distances by means of passive diffusion, but instead must rely on the cell’s machinery for directed active transport involving the use of the cytoskeleton and transport vesicles. Viral capsids and viruses packaged in transport vesicles attach to dynein (and kinesin) motors and move along microtubules to the nucleus. The viruses first move along actin fibers and then along microtubules to reach their sites of replication, and once replicated and assembled move again along the cell’s active transport highway to reach the cell periphery.
18.2Viruses Enter and Exit the Nucleus in Several Ways
Viruses that replicate in the nucleus must pass through the nuclear pore complex, first to get into the nucleus and then again to leave. In some cases, the viruses travel back and forth, in and out of the nucleus several times. Viruses exploit several mechanisms for passage through the nuclear pore

18.3 Ways that Viruses Exit a Cell |
443 |
complex, their choice depending on the virus’ type and size. Virus particles of diameter less than 30 to 40 nm are small enough to pass through the NPC intact, without dissolution of the capsid, but larger viruses cannot pass through without some rearrangement and capsid disassembly. These larger viruses must either wait for dissolution of the nuclear envelope during mitosis, or they must at least partially disassemble to pass through the NPC during interphase.
Proteins that are actively transported into the nucleus are referred to as being karyophilic. These proteins rely on importins and Ran GTPases for passage through the NPC in the manner discussed in Chapter 11. These proteins have nuclear localization signals that interact with importins a and b. Many viruses enter and leave the nucleus in the same manner as cellular proteins. In the case of small viruses that can pass through the NPC intact, the capsids contain proteins bearing nuclear localization signals (NLSs).
Herpes simplex virus Type 1 (HSV-1) is an example of a virus that is too large to pass into the nucleus without disassembly. HSV-1 is an enveloped virus that fuses with the plasma membrane of the host cell. The intact capsid is released into the cytoplasm and is transported to the nucleus where it docks. In order to gain entry to the nucleoplasm of its nondividing host it expels its DNA directly into the nucleoplasm in a spring-loaded-like fashion, leaving its icosahedral 125-nm wide capsid outside the nucleus.
This is not the only mechanism utilized by large viruses to get their DNA into the nucleus. In the case of adenoviruses such as Ad2, the capsids are first disassembled and then the DNA is transported through the NPC into the nucleus. This is accomplished with the use of NLSs resident on proteins that chaperone the DNA through the NPC. Viruses such as HIV-1 utilize yet another mechanism. These viruses are able to bind directly to the nuclear pore, and their DNA can enter the nucleus without requiring the assistance of importins.
18.3 Ways that Viruses Exit a Cell
Viruses forms buds in endosomal vesicles that are transported to the plasma membrane. Viruses are able to exit a cell in at least two different ways. They may exit one at a time through membrane fusion, or they may egress in mass through cell lysis. They also may leave by some other less common means. Enveloped viruses acquire their envelopes from the host cells by budding. They may bud off of an internal membrane or generate a bud from the plasma membrane. These viruses typically exit the cell through membrane fusion where one virus at a time fuses with the plasma membrane and releases the virus into the extracellular spaces. The second common means for release, cell lysis, is typically associated with nonenveloped viruses. These viruses are released in large numbers all at once in a process that causes the rupture of the plasma membrane and death of the host cell.

444 18. Regulation by Viruses
In order to exit the cell, virus particles interact with the cellular machinery responsible for multivesicular body (MVB) biogeneisis. MVBs are a type of endosome having a multivesicular appearance; they play a role in receptor internalization, protein sorting operations, and viral budding. In their role as devices for internalization and protein sorting, the MVBs transport receptors from the plasma membrane to lysosomes for either degradation or recycling back to the plasma membrane. These transport and sorting operations are referred to as the MVB sorting pathway.
The proteins that participate in the vascular protein-sorting in MVBs are called class E Vps’s. The reason for this name is that, in the absence of vascular protein-sorting (Vps) proteins, abnormal endosomes are formed called class E compartments. The class E Vps proteins form complexes called endosomal-sorting complexes required for transport (ESCRTs). In humans, there are four distinct ESCRT complexes. Members of these complexes interact with the exiting viruses through late (L) domains. Late domains are found in virus gag proteins in some viruses and in matrix proteins in others (virus gag and matrix proteins will be discussed shortly). These L domains mediate interactions with the ESCRTs. The motifs found in L domains are protein-protein interaction domains. The three most common binding motifs are PXXP, PPXY, and YXXL. The first of these binds to proteins with SH3 domains; the second binds proteins possessing WW domains, and the third provides attachment for clathrin-associated adapter proteins. The last named motif is often found in cytoplasmic tails of proteins involved in sorting and shipping.
18.4 Viruses Produce a Variety of Disorders in Humans
Viruses are responsible for colds and infections, immunodeficiency disorders, and cancers. Viruses that infect humans include both DNA and RNA viruses. One of the main families of DNA viruses infecting humans is the herpesvirus (Herpes simplex virus, or HHV) family. This family of viruses is a large one and includes the Epstein-Barr virus (HHV-4) and Kaposi’s sarcoma associated herpes (HHV-8). Another common DNA family is the adenovirus family responsible for producing respiratory infections, and still another is the poxvirus family (vaccinia) responsible for smallpox and cowpox. Several families of RNA viruses cause diseases in humans, among which are colds, hepatitis, influenza, measles, mumps, polio, rabies, and AIDS. Members of the most prominent families of viruses and the diseases they cause are listed in Table 18.1. All of these viruses have been studied extensively in the laboratory along with viruses that infect bacteria (bacteriophages), plants, and arthropods (e.g., baculoviruses).
Viruses differ from bacteria in the way they damage hosts: Bacteria release toxins while viruses kill their host cells when they escape from them. In spite of differences in damage-causing mechanisms, both kinds of

18.5 Virus–Host Interactions Underlie Virus Survival and Proliferation |
445 |
TABLE 18.1. Viruses and some of the diseases they cause: Abbreviations—Reverse transcribing (RT); single-strand negative sense (-ss); single-strand positive sense (+ss); double strand (ds).
Virus family |
Type of virus |
Diseases |
Adenovirus |
DNA |
Respiratory infections |
Herpesvirus |
DNA |
Chickenpox, cold sores, encephalitis, genital |
|
|
sores, Kaposi’s sarcoma, mononucleosis, |
|
|
roseola, shingles |
Papovavirus |
DNA |
Warts |
Poxvirus |
DNA |
Cowpox, smallpox |
Calicivirus |
+ssRNA |
Norwalk |
Filovirus |
-ssRNA |
Ebola, Marburg |
Flavivirus |
+ssRNA |
Yellow fever, hepatitis C |
Orthomyxovirus |
-ssRNA |
Influenza A and B |
Paramyxovirus |
-ssRNA |
Measles, mumps, respiratory, colds (children), |
|
|
pneumonia (adults) |
Picornovirus |
+ssRNA |
Polio, hepatitis A, common cold |
Rhabdovirus |
-ssRNA |
Rabies |
Rheovirus |
dsRNA |
Intestinal infections, respiratory infections |
Togavirus |
+ssRNA |
Rubella |
Lentivirus |
RT RNA |
AIDS |
Oncornavirus |
RT RNA |
Human T cell leukemia |
|
|
|
pathogens utilize many of the same strategies to evade detection and suborn host defenses. Like bacteria, viral genomes contain both structural and regulatory genes. The genomes of viruses encode proteins comprising their caspsids, envelopes, and other components. The genes encoding these structural proteins are referred to as structural genes. In addition, viral genomes contain a number of nonstructural genes. These genes encode proteins involved in replication and proteins of a regulatory nature.
18.5Virus–Host Interactions Underlie Virus Survival and Proliferation
Viruses interact with their hosts in order to survive. They manipulate elements of the host control layer in order to create environments that support replication and proliferation. They turn off antiviral activities, prevent apoptosis, and alter the cell cycle progression. Shown in Table 18.2 are some examples of eukaryotic control proteins targeted by viruses. One of the groups of signaling proteins targeted by viruses is the cytokine group that coordinates the activities of leukocytes. The cytokine signaling events are disrupted by the viruses, thereby reducing the effectiveness of the immune system in responding to the viral invasion.
Another group of signaling elements targeted by the viruses are the antigen-presenting elements that present peptides on their surface derived

446 18. Regulation by Viruses
TABLE 18.2. Eukaryotic cell components targeted by some well-known viruses.
Control layer elements |
Viruses (and signaling elements interfered with) |
Antigen-presenting elements |
Adenovirus (MHC class I, II), Epstein-Barr virus (MHC, |
|
class II) |
Apoptosis system |
Adenovirus (Fas, Bcl-2, caspases), baculovirus (caspases), |
|
cowpox (Fas, caspases), Epstein-Barr virus (Bcl-2), |
|
vaccinia (caspases) |
Cytokine signaling |
Cowpox (TNF), human herpesvirus-8 (CC and CXC |
|
chemokines and their receptors), vaccinia (IL-1b, |
|
IFNa/b) |
Interferon signaling |
Adenovirus (Jak/STAT), baculovirus (PKR), influenza virus |
|
(PKR), poliovirus (PKR), reovirus (dsRNA) |
|
|
from the viruses of the invaded cells. A third set of targets is the proteins that regulate apoptosis. Cells infected by viruses are killed and induced to undergo apoptosis. The viruses manipulate this protective response; in some cases they turn off apoptosis and in others they use the process to hitch a ride out of the cell. Finally, receptors bound in the plasma membrane are targeted by viruses in order to gain entry into the cell.
18.6Multilayered Defenses Are Balanced by Multilayered Attacks
Pathogens such as viruses and bacteria enter a body, or host, establish themselves in a specific locale, or niche, multiply, cause damage, and exit. Host defenses are mounted at multiple levels. The outermost layer is the erection of mechanical barriers such as the skin, and chemical barriers such as antibacterial defensins. Next, the innate immune response is triggered and then the adaptive immune response is.
The approach used by viruses to interfere with the host’s control system is a multilayered one. Viruses, for example, overcome the acquired immunity response (antigen-presenting elements) by interdicting expression of the MHC Class I and II products at the transcription level at the translation level, by forcing the retention of the surface molecules in the cytoplasm, by accelerating their removal from the surface, and by stepping up their deactivation.
The survival of a pathogen in a host is critically dependent on its ability to deal with an arsenal of host defenses. Viruses and interferon systems have coevolved. One striking consequence of this coevolution is that practically all viruses have developed some level of resistance to the interferon system. The counterattacks take several forms and are multileveled, like the response to acquired immunity. Depending on the specific virus and target, the virus may

18.8 Hepatitis C Virus Disables Host Cell’s Interferon System |
447 |
•block the production of interferons,
•supply interferon decoys,
•block interferon signaling, and
•block the ability of the interferon signaling targets to properly act.
At the most fundamental level, what is happening is that both host and pathogen (viral and bacterial) have coevolved. Both have developed control layers tuned to the other, promoting adaptation and evolution of each. The overall result is a balance, an equilibrium situation of coexistence of host and bacterial and viral pathogens.
18.7 Viruses Target TNF Family of Cytokines
The TNF family of cytokines and their receptors, and the apoptosis machinery, are targeted by viruses. The TNF family of cytokines and their receptors are central signaling elements of the adaptive immune response. Members of this family regulate secondary lymphoid organ development, lymph node formation, and apoptosis signaling. A variety of viruses including hepatitis C virus, Epstein-Barr virus, adenoviruses, and poxviruses elude destruction by the adaptive immune response by expressing proteins that alter the operation of the TNF machinery.
The hepatitis C virus core protein is a structural gene product that is able to bind to and modulate TNF family receptors. In response to these binding events cytokine signaling is impaired and cytotoxic T lymphocytes do not adequately respond to the viral invasion. Adenoviruses target the cellular apoptosis machinery triggered by the TNF family member Fas (Table 18.2). The viruses are able to induce the internalization of this receptor, thereby turning off the ability of the cells to kill the virus. Adenoviruses use other mechanisms as well to elude the adaptive immune response. For instance, they interfere with antigen presentation and T cell recognition. Each of the adenoviral evasion tactics utilizes a different subset of viral gene products.
Cells infected with a virus are usually induced to undergo apoptosis in order to prevent viral replication. Interestingly, some viruses have evolved ways to use this mechanism to their own advantage. They induce cell death at the end of their replication cycle with the result that they are encapsulated in a protective package that is taken up by neighboring cells. By this means they spread and elude detection at the same time.
18.8Hepatitis C Virus Disables Host Cell’s Interferon System
The hepatitis C virus is believed to infect more than 2% of the world’s population. The virus causes chronic hepatitis and can lead to cirrhosis of the liver and/or liver cancer. The virus is a member of the Flaviviridae

448 18. Regulation by Viruses
FIGURE 18.1. Organization of the genome of the hepatitis C virus: Proteins shown as dark-shaded boxes are structural (S) in character while those in light-shaded boxes are nonstructural (NS). From left to right the proteins are: C—nucleocapsid protein, sometimes called the “core protein”; E1 and E2—glycoproteins belonging to the viral envelope; p7—linker protein of unknown function; NS2 through NS5B— NS enzymes. Arrows denote cleavage sites. Solid arrows indicate sites cleaved by host cellular signalases; the dashed arrow indicates the site cleaved by NS2, and the dotted arrows denote sites cleaved by NS3.
family of viruses, which includes the yellow fever virus and several forms of encephalitis-inducing virus.
The hepatitis C virus (HCV) contains a single strand positive sense RNA (+ssRNA) molecule approximately 9.6 kBp in length that functions as a messenger RNA. The genome encodes a single open reading frame that is preceded at the 5¢ end by a noncoding region and terminated at the 3¢ end by another such region. The resulting polypeptide chain, or polyprotein, is proteolytically processed during and after translation into 10 proteins (Figure 18.1). The four amino-terminal-most are structural in character. The first protein, C, encodes the capsid protein, and the next two, E1 and E2, are glycoproteins that are inserted in the viral envelope. It is customary to refer to proteins forming the virus capsid and envelope as “structural” proteins and proteins involved in replication and signaling/regulation as “nonstructural.” The fourth structural protein, p7, is a linker that separates the structural from nonstructural proteins.
The nonstructural genes encode several different kinds of enzymes. The NS5B gene encodes an RNA-dependent RNA polymerase that is responsible for viral replication. The NS2 and NS3 genes encode proteases that, along with cellular proteases, chop up the large polyprotein into the structural and nonstructural proteins at the sites indicated in Figure 18.1. The NS4B and NS5A gene products are regulatory in nature. The regulatory gene products along with the E2 glycoprotein target the interferon system of antiviral defense, allowing the viruses to survive and proliferate even when that system is activated.
Recall from Chapter 9 that the cell’s first line of defense to a viral attack is the interferon system. During viral replication dsRNA is produced and the detection of these molecules alerts the cell that viruses are present. A sensor of dsRNA called protein kinase R (PKR) activates the interferon response. PKR shuts off translation and stimulates transcription of a host

18.9 Human T Lymphotropic Virus Type 1 Can Cause Cancer |
449 |
of antiviral factors. Interferons are produced that signal to other cells through interferon receptors and Jak/Stat proteins to generate a global response to the viral invasion. The favored treatment for patients with hepatitis C is IFNa, or a-interferon. However, this form of treatment, the best available at present, is ineffective in more than half of patients. In these many cases, the virus is able to overcome the antiviral activities of the interferons.
The interferon line of defense is breached by hepatitis C virus through its NS5A, NS4B, and E2 proteins, which inhibit the activity of PKR. The NS5A protein inhibits not only the PKR protein but also the IRF1 transcription factor, which acts downstream of PKR to trigger transcription of antiviral genes from the ISRE promoter. The E2, NS5A, and NS4B proteins enhance translation from what is termed an HCV internal ribosomal entry site, or IRES. The net effect of E2, NS4B, and NS5A is to throttle back expression of antiviral genes and maintain the translation capabilities of the host cell so that the viruses can replicate.
18.9Human T Lymphotropic Virus Type 1 Can Cause Cancer
Certain retroviruses can induce the transformation of normal cells into cancerous ones. One group of retroviruses that can trigger cancers is the oncogenic (oncornaviruses) viruses, typified by the Rous sarcoma virus. This virus has picked up an oncogene from a host some time in its past, then deliveres it to its new hosts when it invades a cell. In the case of Rous sarcoma virus, the oncoprotein is Src, a tyrosine kinase involved in the growth pathway. This protein was discussed in several places in the text. To distinguish cellular from viral forms, one is designated as “c-Src” and the other as “v-Src.” The v-Src oncoprotein is of considerable historical importance, as it was the first oncoprotein to be discovered.
A different mechanism is used to transform cells by viruses such as bovine leukemia virus (BLV) and human T lymphotropic virus type 1 (HTLV-1). The latter is associated with T cell leukemia. These retroviruses do not carry oncogenes. Instead regulatory proteins are encoded in their genomes, and these proteins along with promiscuous, virally supplied promoters that accompany them fatally alter the operation of the cellular control layer. Viruses belonging to this group cause a variety of cancers in livestock, while HTLV-1 and -2 promote cancer (adult T cell leukemia) in humans.
The organization and placement of control sites is an essential feature of a genome. Retorviruses bring their own control sites with them. These control sites are known as long terminal repeats, or LTRs. There is an LTR at the 5¢ end and the other at the 3¢ end. The one at the 5¢ end is used for transcription control. Each LTR is composed of three regions: a U5 region, an R region, and a U5 region. The U3 region controls viral transcription
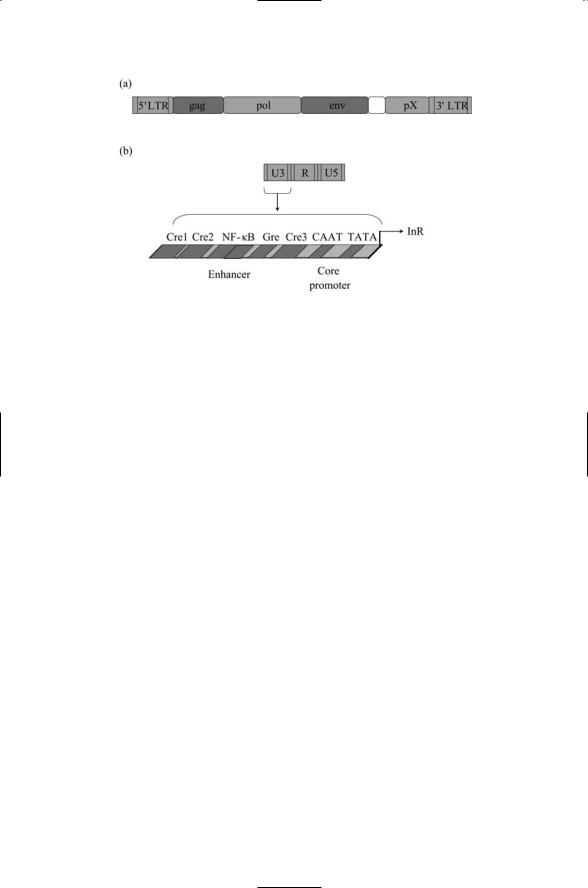
450 18. Regulation by Viruses
FIGURE 18.2. Retroviral control region: (a) A minimal retrovirus, containing gag, pol, and env genes, is shown bounded at both ends by long terminal repeats (LTRs) and a pX region near the 3¢ end. (b) An expanded view of the 5¢ LTR showing its U3, R, and U5 regions—A further expanded view of the U3 region depicts a canonical set of DNA binding sites (responsive elements) for transcription factors. Abbreviations: cyclic AMP responsive element (Cre); glucocorticoid responsive element (Gre); initiation, or start, site for RNA transcription (InR), located at the boundary between U3 and R.
while the R and U5 regions are essential for viral replication. As illustrated schematically in Figure 18.2, the U3 region is endowed with binding sites for many of the key families of eukaryotic transcription factors. These sites are capable of binding to both viral and cellular regulatory proteins.
A unique feature of HTLV-1 is the presence near the 3¢ end of its genome of a region called pX that codes for several regulatory proteins. One of these proteins, called Tax, binds to CREB proteins at the various Cre sites not only in the viral genome but also in the cellular genome. Tax interacts with a variety of cellular regulators of transcription including not only CREB and cofactors but also NF-kB. Through its interactions with the cellular regulators, Tax changes the regulatory decisions of the T cell’s control circuitry, immortalizing and transforming the cells.
18.10 DNA and RNA Viruses that Can Cause Cancer
Listed in Table 18.3 are a number of viruses that are known to transform cells in humans. The list includes both DNA and RNA viruses. The HTLV- 1 virus appears in the table as a member of the retrovirus group that promotes T cell leukemias in adults. One of the recent additions to the list of viruses known to promote cancer is Kaposi’s sarcoma-associated her-