

Глава 3. Сократительные компоненты мышцы
Кроме того, эластичность соединительных тканей сухожилий, которые прикрепляют концы мышцы к кости, восстанавливает исходную длину мышцы.
Молекулярная (химическая) основа расслабления. Химические реакции, связанные с расслаблением, недостаточно хорошо изучены. По мнению большинства ученых, расслабление обусловлено прекращением процесса сокращения. Во время расслабления сочетания каль-ций-тропонин отделяются и ионы кальция возвращаются в мешочки саркоплазматического ретикулума. Поскольку тропонин уже больше не связан с кальцием, он предотвращает взаимодействие актина с миозином. Это способствует диссоциации актина и миозина и «повторному скольжению» филаментов назад в положение покоя. Иными словами, сокращение «включается» выделением кальция и «выключается» его устранением.
ТЕОРИЯ МЫШЕЧНОГО УДЛИНЕНИЯ
Мышечные волокна не способны сами по себе удлиняться или растягиваться. Чтобы произошло их удлинение, к мышце должна быть приложена сила извне. Это может быть сила притяжения, сила движения, сила мышц-антагонистов, действующих на противоположную сторону сустава, сила, прилагаемая другим человеком или другой частью своего тела.
Теоретическое ограничение реакции на растяжение сократительного компонента мышечной клетки, основанное на теории скольжения филаментов, можно определить путем измерения под микроскопом длины сар-комера, миозиновых филаментов, актиновых филаментов и Н-зоны. Рассмотрим, как изменяется длина саркомера при растяжении по сравнению с длиной в покое (табл. 3.1).
Таблица 3.1. Длина сократительных компонентов в покое
|
Саркомер |
2,30 мкм |
|
Миозин |
1,50 мкм |
|
Актин |
2,00 мкм |
|
Н-зона |
0,30 мкм |
При максимальном растягивании саркомера до точки разрыва его длина может достигнуть приблизительно 3,60 мкм. Разрыв саркомера, естественно, нежелателен. Наша главная задача — растянуть саркомер до такой длины, при которой происходит незначительное перекрывание филаментов и хотя бы один поперечный мостик сохраняется между филаментами актина и миозина. Эта длина составляет около 3,50 мкм. Таким образом, при длине саркомера в покое 2,30 мкм его сократительный компонент способен увеличиться на 1,20 мкм, то есть увеличение по сравнению с состоянием покоя составляет свыше 50 %. Если длина саркомера в покое 2,10 мкм, а все остальные факторы остаются постоянными, сократительный компонент мышцы может увеличить свою длину по сравнению с длиной в покое на 67 %. Такая растяжимость дает возможность нашим мышцам двигаться с большой амплитудой.
39
Наука о гибкости
ТЕОРИЯ СКОЛЬЖЕНИЯ ФИЛАМЕНТОВ
В
соответствии с первоначальной теорией
скольжения филаментов толстый и тонкий
филаменты попросту скользят относительно
друг друга при
растягивании саркомера. Во время такого
изменения длины саркомера длина
толстого и тонкого филаментов не
изменяется, но степень их взаимного
перекрытия становится меньше (см. рис.
3.10). Однако мы знаем, что механизм,
посредством которого происходит
удлинение саркомера, является
более сложным, чем первоначально
предполагалось. Например, что-то должно
в конечном итоге предотвращать
чрезмерное растяжение саркомера.
Эту защитную функцию выполняет
соединительный филамент.
Результаты
предыдущих структурных исследований
показали, что каждый филамент титина
в саркомере состоит из двух сегментов.
Сегмент между
Z-линией
и краем А-диска, как было обнаружено,
является достаточно
растяжимым. В то же время другой сегмент,
который взаимно перекрывается
толстыми филаментами, является
тугоподвижным и нерастяжимым. На
основании этих открытий было предложено
несколько рабочих моделей. В модели
Поллака (1990; рис. 3.11) предполагается,
что соединительный
филамент абсорбирует растяжение в
двух своих элементах: тро-помиозине
и титине. Первоначально растягивание
происходит легко (рис. 3.11, а,
б).
Во время этого
незначительного растягивания тропомиозин
рекрис-таллизируется и сообщает слабое
статическое усилие саркомеру. На этом
этапе титин не
растягивает-
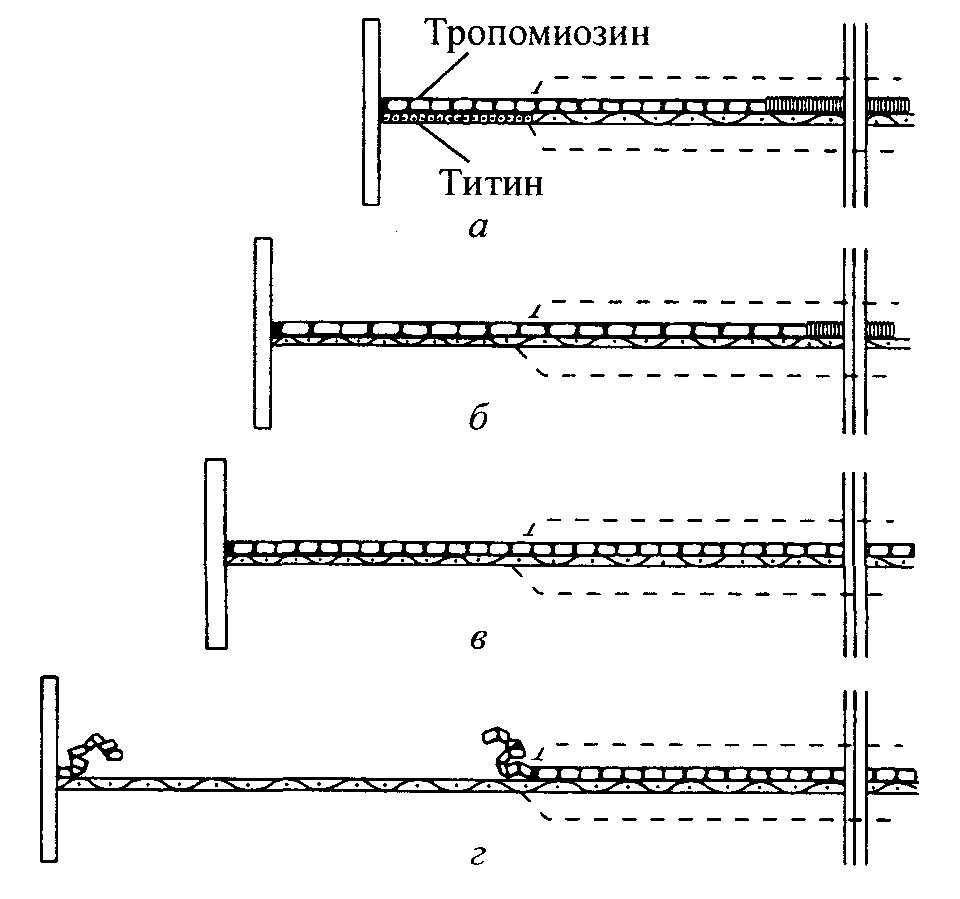
ся настолько, чтобы ощущалась возвратная сила. Таким образом, саркомер растягивается без особого труда. По мере продолжения растягивания происходит кристаллизация всего тропо-миозина. Вследствие этого тропомиозиновая нить становится нерастяжимой и дальнейшее растяжение саркомера невозможным. Таким образом, считают, что именно тропомиозин обусловливает «рабочую» длину саркомера (рис. 3.11, в).
Однако даже эта система имеет свои пределы. При продолжении удлинения тропомиозиновая нить рвется (рис. 3.11, г). Разрыв тропо-миозиновой нити не означает разрушение саркомера.
40
Г л а в а 3 ■ Сократительные компоненты
мышцы

Длина саркомера, мкм
Рис. 3.12. Сегментное вытяжение филаментов титина как структурная основа кривых
нагрузки-напряжения мышц в покое. Показаны предполагаемые структурные явления,
происходящие в филаментах титина, лежащие в основе кривых нагрузки-напряжения
(Wang, McCarter, Wright, Beverly and Ramirez-Mitchell, 1991)
Специальная «дублирующая» система готова сохранить целостность саркомера во время «ремонта» титиновой нити (рис. 3.11, г).
Вскоре после появления модели Поллака в 1990 г. Ванг с коллегами (1991) предложили свою модель. В соответствии с этой моделью по мере растяжения саркомера растяжимый сегмент титина перемещается в результате удлинения. Чем выше степень удлинения, тем более длинным становится растяжимый сегмент вследствие рекрутирования ранее нерастяжимого титина, когда его соединение с толстыми филаментами начинает проскальзывать или когда происходит деформация дистальных концов толстых филаментов (рис. 3.12). Филамент титина, который простирается от Z-линии к М-линии, имеет два механически обособленных сегмента: растяжимый сегмент в I-диске (незаштрихован) и нерастяжимый сегмент, ограниченный взаимодействием с толстыми филаментами (заштрихован). При сокращенной длине саркомера (SLo) титин может быть «дряблым», и растяжение до SLe не приводит к изменению длины контура или производству значительного усилия.
41
Наука о гибкости
За пределами SL, линейное вытяжение титанового сегмента приводит к экспоненциальному увеличению напряжения. При SL, растяжимый сегмент становится длиннее вследствие рекрутирования ранее нерастяжимого титина после начала его открепления от толстых филаментов или деформации дис-тальных концов толстых филантов, ведущей к выравниванию напряжения. Предполагают, что чистая контурная длина растяжимого сегмента титина длиннее в саркомере, выражающем более крупную изоформу титина. Вследствие этого SLe и SL увеличиваются, а кривые нагрузки-напряжения различных мышц можно нанести на график как функцию напряжения растяжимого сегмента титина (ТЕ/ТЕ ). Дальнейшее удлинение приводит к разрыву титановой нити. В сущности, тропомиозин можно сравнить с поездом, а титин с рельсами. Когда железнодорожный поезд (т. е. тропомиозин) сходит с рельс, не все оказывается потерянным. Его просто нужно снова поставить на рельсы. Если же повреждаются рельсы, то ничего уже нельзя сделать. Таким образом, именно титин обеспечивает целостность саркомера.
ДРУГИЕ ФАКТОРЫ, ОГРАНИЧИВАЮЩИЕ ДИАПАЗОН ДВИЖЕНИЯ
Ряд факторов могут ограничивать диапазон движения сустава. Это может быть мышечный дисбаланс, неадекватный мышечный контроль, возраст мышцы или ее иммобилизация.
Мышечный дисбаланс. Здоровые мышцы сохраняют структурный гомеостаз. Ключом такого структурного баланса является взаимная тяга мышц-антагонистов, расположенных на противоположной стороне сустава (рис. 3.13). Дисбаланс этих сил тяги может повлиять на диапазон движения. Мышечный дисбаланс может быть обусловлен рядом факторов, включая наличие гипертонических мышц (т. е. мышц, находящихся в состоянии контрактуры или спазма) или слабых мышц.
Неадекватный мышечный контроль. Даже если человек от рождения наделен достаточной гибкостью, локальный мышечный контроль может быть неадекватным для выполнения определенных элементов, требующих проявления гибкости. Это объясняется тем, что многие из них состоят из дополнительных компонентов. В данном контексте мы рассматриваем мышечный контроль как наличие адекватного баланса, координации или контроля за частью (частями) тела или достаточной силы для выполнения конкретного элемента, требующего проявления гибкости. Например, чтобы выполнить такой элемент, как «ласточка», требуется умение сохранять равновесие. Кроме того, необходимо обладать достаточной силой, чтобы принять и сохранить необходимое положение. Выполнение более сложных двигательных умений требует соответствующего сочетания всех необходимых качеств и компонентов, подготовленности.
Влияние процесса старения на мышцы. Естественный процесс старения ведет к снижению функциональных возможностей мышцы. Если человек ведет малоподвижный образ жизни, страдает какими-нибудь заболеваниями и т. д., этот процесс протекает намного быстрее. С физиологической
42

Г л а в а 3 ■ Сократительные компоненты
мышцы

Рис. 3.13. Мышечный баланс: а — мышечный баланс обеспечивает структурный баланс;
б — дисбаланс нередко наблюдается при ослаблении одной мышцы; в — иногда дисбаланс
обусловлен тем, что одна из мышц оказывается слишком сильной (Walther, 1981)
точки зрения наиболее очевидным дегенеративным изменением, связанным с процессом старения, является постепенная атрофия мышечной массы. Она обусловлена снижением размера и количества мышечных волокон (Grab, 1983; E.Gutmann, 1977; Hooper, 1981). Как замечает Уилмор (1991), «... это снижение количества саркомеров может обусловливать снижение подвижности, обычно наблюдаемое у пожилых людей». Отмечаются значительные колебания возраста, в котором проявляются эти изменения, существенно варьируется и степень изменений. С возрастом также сокращается количество нервных клеток в скелетно-мышечной системе (E.Gutmann, 1977).
По мере атрофии мышечных волокон происходит их замена жировой и волокнистой (коллагеновой) тканью. Коллаген — главный компонент соединительной ткани — характеризуется исключительно низкой эластичностью. Поэтому вполне очевидно, что даже незначительное увеличение количества коллагена в мышце приводит к существенному снижению ее подвижности. Это явление изучали Альнакиб, Аль-Заид и Голд-спинк (1984), использовав для этого камбаловидную мышцу и длинный разгибатель пальцев крыс. Их исследования подтвердили, что общее содержание коллагена с возрастом постоянно увеличивается. Кроме того, они обнаружили в молодой мышце более низкую интенсивность развития пассивного напряжения на каждую единицу увеличения длины. В мышце взрослого животного пассивное напряжение развивалось с большей интенсивностью.
«Данные, касающиеся тугоподвижности мышц и соединительной ткани, характеризуются корреляцией, за исключением данных о камбаловидной мышце старых животных. Пассивное механическое поведение мышцы непосредственно связано с концентрацией коллагена».
Таким образом, эти изменения в определенной степени обусловливают возрастное снижение гибкости.
Влияние иммобилизации. Способность мышцы адаптировать свою длину в зависимости от положения продемонстрировал в экспериментах
43
Наука о гибкости
еще Марви (Marvey, 1887). В последние 20 лет механизмы адаптации длины мышцы изучали как на клеточном, так и на ультраструктурном уровнях. Исследования, проведенные Голдспинком (1968, 1976) и П.Уильямсом и Голдспинком (1971), показали, что увеличение длины мышечного волокна во время нормального развития связано со значительным увеличением количества саркомеров вдоль длины волокон. Так как длина филаментов актина и миозина постоянна, адаптация взрослой мышцы к различной функциональной длине, по-видимому, должна включать производство или удаление определенного количества саркомеров с тем, чтобы сохранить правильную длину саркомера относительно всей мышцы (Goldspink, 1976; Tabary и др., 1972).
При иммобилизации камбаловидной мышцы взрослой кошки в удлиненном положении мышца адаптируется к новой длине. Табари с коллегами (1972) обнаружили, что это удлинение осуществляется за счет появления около 20 % новых саркомеров. П.Е.Уильямс и Голдспинк (1973) обнаружили, что новые саркомеры добавляются на концах имеющихся мио-фибрилл. В случае денервации и иммобилизации в удлиненном состоянии производится около 25 % саркомеров (Goldspink и др., 1974). После прекращения иммобилизации «нормальная» и денервированная мышца быстро восстанавливают свою первоначальную длину (Goldspink и др., 1974; Tabary и др., 1972). Совсем недавно было проведено исследование с целью определить, способна ли внеглазная мышечная система трех обезьян адаптироваться таким образом, как мышцы конечности кошек (A.B.Scott, 1994). Исследования показали увеличение длины глазных мышц на 18, 25 и 33 % вследствие наложения швов.
При иммобилизации конечности с мышцей в сокращенном положении мышечные волокна теряют до 40 % саркомеров (Tabary и др., 1972). При денервации и иммобилизации в сокращенном положении наблюдали снижение числа саркомеров на 35 % (Goldspink и др., 1974). Однако затем количество саркомеров восстанавливается по мере восстановления длины мышцы (Goldspink и др., 1974; Tabary и др., 1972).
Таким образом, адаптация количества саркомеров в зависимости от функциональной длины мышц происходит не под нейронным контролем. Скорее всего, реакция на количество пассивного напряжения, которому подвергается мышца, носит миогенный характер (Goldspink, 1976; P.E.Williams, Goldspink, 1976).
Наряду со снижением длины волокон, а также количества и длины саркомеров ученые выявили и уменьшение растяжимости (увеличение пассивного сопротивления) мышц, иммобилизованных в сокращенном положении (Goldspink, 1976; Goldspink, Williams, 1979). Этот процесс происходил независимо от того, была ли мышца денервирована или нет (Goldspink, 1976). Голдспинк и Уильяме (1979) также обнаружили, что потеря соединительной ткани происходит с меньшей интенсивностью, чем потеря сократительной ткани мышцы. Таким образом, относительное количество соединительной ткани увеличивается (Goldspink, 1976; Goldspink, Williams, 1979). Кроме того, П.Е. Уильяме и Голдспинк (1984) установили, что волокна коллагена в иммобилизованной мышце распола-
44
Г л а в а 3 ■
Сократительные компоненты мышцы
гаются
под более острым углом к оси мышечных
волокон, чем в обычной мышце.
Это изменение, очевидно, влияет на
эластичность мышцы.
Снижение степени растяжимости, вероятно, является защитным механизмом, который не допускает резкого перерастяжения мышцы (Goldspink, 1976; Goldspink, Williams, 1979). Этот механизм особенно важен для сокращенной мышцы (т. е. мышцы, утратившей саркомеры), поскольку растягивание даже в обычном диапазоне движения приведет к перерастяжению сарко-меров и филаменты миозина и актина не смогут осуществить взаимное перекрытие, что вызовет временное повреждение мышцы (Goldspink, 1976; Tabary и др., 1972). С другой стороны, изменения в эластичных свойствах мышцы, иммобилизованной в удлиненном положении, не произойдут, поскольку адаптация обратно пропорциональна, а вероятность перерастяжения мышцы такая же, как и для обычной мышцы (Tabary и др., 1972).
Однако указанное выше снижение степени растяжимости не является просто защитной функцией. Главным последствием изменений количества саркомеров и длины мышцы является смещение кривой длины-напряжения мышцы влево (при иммобилизации мышцы в сокращенном положении) или вправо (при иммобилизации в удлиненном положении). Эти изменения длины помогают мышце адаптироваться к генерированию оптимальных уровней напряжения при новом положении и новой длине.
На основании приведенных выше результатов П.Е.Уильямс с коллегами (1988) решили определить, обусловливает ли недостаточное растяжение или недостаточная сократительная активность сокращение количества саркомеров, увеличение концентрации коллагена и увеличение тугопод-вижности мышц, которое наблюдается при их иммобилизации в сокращенном положении. Они обнаружили, что аккумуляция соединительной ткани, имеющая место в иммобилизованных мышцах, может быть предотвращена либо путем пассивного растяжения, либо путем активной (электрической) стимуляции.
В связи с этим возникает вопрос, являются ли короткие периоды растягивания достаточно эффективными для предотвращения изменений в соединительной ткани мышц, длине волокон и количестве саркомеров, т. е. способствуют ли они сохранению диапазона движений сустава? Исследование, проведенное П.Е. Уильямсом (1988) на мышах, показало, что периоды пассивного растягивания продолжительностью всего 15 мин (через день) обеспечивают нормальные пропорции соединительной ткани. В то же время они не способствуют предотвращению снижения длины мышечных волокон, которое ведет к заметному ограничению диапазона движения.
МЕХАНИЗМ ПАССИВНОГО РАСТЯГИВАНИЯ В МИОФИБРИЛЛОГЕНЕЗЕ
Из предыдущего раздела мы узнали, что иммобилизация мышцы в удлиненном положении ведет к увеличению длины мышечных волокон, которое, в свою очередь, связано с увеличением числа саркомеров в мио-
45
Наука о гибкости

Рис. 3.14. Распределение мРНК у мышечно-сухожильного соединения растянутых скелетных волокон (Russell, Dix Haller and Jacobs-El, 1992)
фибриллах и вдоль волокон. Специфический участок вновь синтезированных саркомеров находится вблизи мышечно-сухожильного соединения. В последние годы ученые обратили внимание на механизм клеточного контроля, лежащий в основе удлинения и гипертрофии мышечных волокон (миофибриллогенез).
Исследования Дикса и Айзенберга (1990, 1991а, 19916) показали, что накопление мРНК медленного окислительного миозина в конце мышцы в растянутых волокнах является более высоким, чем в контрольных. «Такое локальное накопление мРНК обеспечивает региональный синтез сократительных белков, быструю «сборку» саркомеров и распространение мио-фибрилл» (Dix, Eisenberg, 1990, 1993). В частности, «большое цитоплазма-тическое пространство, в котором содержатся полисомы, «открывается» между миофибриллами и сарколеммами мышечно-сухожильного соединения удлиняющихся волокон, и встречается много формирующихся мио-фибрилл» (B.Russell, Dix, Haller, Jacobs-El, 1992; рис. 3.14). Миозин мРНК (X) накапливается на концах растянутых волокон в увеличенном цитоплаз-матическом пространстве между сарколеммами и миофибриллами (тонкие и толстые филаменты). Эта мРНК способствует локальному синтезу сократительных белков и миофибриллогенезу. Ядра зачастую расположены проксимально к концам этих удлиненных волокон. Растянутые мышцы также удлиняются вследствие появления пролиферирующихся миотрубо-чек (более мелкие клетки справа), которые впоследствии могут слиться с существующими волокнами. Кроме того, растянутые мышцы также удлиняются в результате появления пролиферирующихся миотрубочек (Moss, Leblond, 1971).
Каким же образом растягивание увеличивает производство мРНК? Известно, что для роста клеток необходимы гормоноподобные молекулы — факторы роста. Возможные механизмы были предложены Б.Расселлом, Диксом, Халлером, Джекобс-Элемом (1992). Однако этот вопрос требует дальнейших исследований.
46