

Глава 3- Сократительные компоненты мышцы
|
Соединительный |
Тонкий |
Толстый |
|
филамент |
филамент |
филамент |
|
(титин) |
(актин) |
(миозин) |
I-мостик А-мостик
— 1-диск»-Ц
М-мостик
А-диск -
43 нм
Z-линия
• 1-диск -
Рис.
3.8. Диаграммное изображение основных
структур саркомера
ПРИНЦИПИАЛЬНАЯ СТРУКТУРА САРКОМЕРА: РАБОЧАЯ МОДЕЛЬ
Схематическая модель принципиальных структур саркомера изображена на рис. 3.8 (Pollack, 1990). Мы уже знаем, что саркомер содержит, по меньшей мере, три продольно ориентированные структуры: актин (тонкие филамен-ты), миозин (толстые филаменты) и титин (соединительные филаменты). Кроме того, саркомер включает еще, по меньшей мере, два поперечных соединения, не только между толстыми филаментами в А-диске (М- и А-мости-ки), но и между соединительными филаментами в I-диске (1-мостики).
Саркотубулярная система

Рис. 3.9. Саркотубулярная система (Fawcett, 1986)
3*
35
Наука о гибкости
сом, она подвергается деполяризации. Одновременно деполяризуется вся Т-система, обеспечивая тем самым передачу электрического импульса всем саркомерам мышечного волокна.
Импульсы после этого передаются рукавообразной системе мешочков и трубочек саркоплазматического ретикулума, где содержатся ионы кальция. После деполяризации Т-системы этот электрический разряд передается на мембрану саркоплазматического ретикулума, увеличивая ее проницаемость. В результате этого ионы кальция покидают мешочки саркоплазматического ретикулума.
Теория сокращения
Функция мышцы состоит в производстве или генерации напряжения. Этот процесс генерации напряжения называется сокращением. Главная цель мышечного сокращения — производство движения. Две другие функции, связанные с сокращением — это сохранение положения и производство телом тепла. После инициации мышечного сокращения происходит целый ряд физических и химических явлений.
Ультраструктурная (физическая) основа сокращения. Механизм, посредством которого мышца сокращается, расслабляется или удлиняется, можно объяснить ультраструктурой саркомера. Наиболее известной теорией является теория скольжения филаментов (рис. 3.10, а-в). В соответствии с этой теорией, изменения длины саркомера обусловлены (исключительно) относительным скольжением толстого и тонкого филаментов. Точный механизм, регулирующий сократительные элементы, еще не совсем хорошо изучен. По мнению Поллака (1983, 1990), например, эта теория не имеет под собой достаточно прочной основы. Не так давно появилась новая гипотеза, согласно которой после периодов сокращения саркомера следуют паузы, во время которых длина не изменяется или изменяется незначительно, в результате чего форма волны сокращения имеет ступенчатый характер (Pollack и др., 1977).
При максимальном сокращении длина саркомера может уменьшиться на 20-50 % по сравнению с его длиной в покое. При пассивном растягивании она может превысить обычную длину на 120 %. Результаты наблюдений показывают, что длина А-дисков и, следовательно, толстых филаментов всегда остается постоянной. Остается постоянным на всех этапах обычного сокращения и расстояние между Z-линией и краем Н-зоны, а это свидетельствует о том, что длина тонких актиновых филаментов также не подвергается изменениям. Исходя из этих наблюдений, ученые пришли к выводу, что изменение длины мышцы обусловлено скольжением толстого и тонкого филаментов относительно друг друга.
Таким образом, когда мышца сокращается, актиновый и миозиновый фи-ламенты скользят один относительно другого, в результате чего каждое волокно сокращается (теория скольжения филаментов). Чтобы этот процесс имел место, Z-линия саркомера должна приблизиться к А-диску, что приведет к постепенному сужению и последующему «устранению» I-дисков и Н-зоны.
36
Г л а в а 3 ■ Сократительные компоненты мышцы

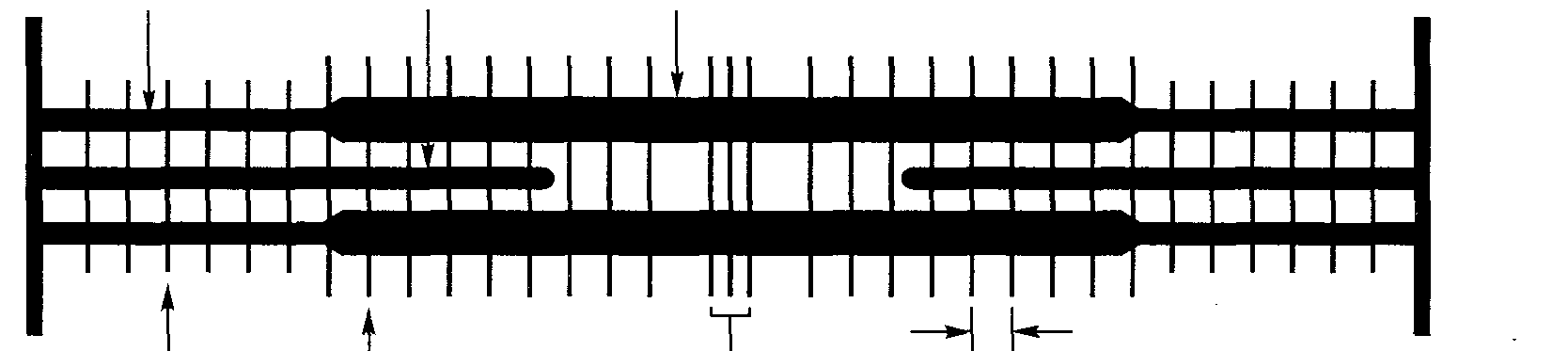
. 3,65 мкм (а + б) -

Интервалы между полосами, мкм в
Рис. ЗЛО. Схематическое изображение изменений тетанического напряжения в интервалах между полосами. Стрелки вверху расположены напротив интервалов, в которых имеет место взаимное перекрывание филаментов (а). Схематическая диаграмма филаментов с указанием номенклатуры соответствующих измерений (б). Критические этапы увеличения степени взаимного перекрытия толстых и тонких филаментов по мере сокращения саркомера (в) (Gordon,Huxley,Julian, 1966)
37
Наука о гибкости
Молекулярная (химическая) основа сокращения. Немедленным источником энергии для осуществления мышечных сокращений служит расщепление аденозинтрифосфата (АТФ), обусловленное нервными импульсами. Когда нервные импульсы поступают в волокно скелетной мышцы, они распространяются по сарколемме и двигаются вовнутрь через Т-трубочки. Это приводит к увеличению проницаемости и вызывает выделение ионов кальция (Са2+) из мешочков саркоплазматического рети-кулума в саркоплазме. Считают, что в состоянии покоя молекулы тропо-миозина находятся сверху активных участков на филаментах актина, что предотвращает привязывание на поперечных мостиках миозина и актино-вом филаменте. После выделения Са2+ они связываются с молекулами тропонина на филаменте актина. Этот процесс называется «включением» активных участков на филаменте актина. Одновременно происходит зарядка незаряженного комплекса поперечного мостика АТФ, что позволяет актину и миозину образовать актомиозиновый комплекс. Это, в свою очередь, активирует ферментный компонент миозинового филамента, который называется миозин АТФ-аза. Миозин АТФ-аза расщепляет АТФ на АДФ и Рн (неорганический фосфат), что сопровождается выделением энергии. В результате выделения энергии изменяется угол поперечных мостиков и они скользят над филаментом миозина к центру саркомера. Мышца сокращается и производит напряжение. Таким образом, очевидно, что активация мышц полностью зависит от нервных импульсов. Без нервного импульса генерирование мышечного напряжения просто невозможно.
ТЕОРИЯ МЫШЕЧНОГО РАССЛАБЛЕНИЯ
Способность мышцы расслабляться — неотъемлемое условие оптимального движения и состояния здоровья. Поэтому процесс мышечного расслабления тщательно изучался как на физическом, так и на химическом уровнях. Вместе с тем точный механизм расслабления еще не до конца изучен. В следующих разделах мы рассмотрим физическую и химическую основу расслабления.
Ультраструктурная (физическая) основа расслабления. Мышечное расслабление является полностью пассивным. Когда в мышечные волокна не поступают нервные импульсы, они расслабляются. Следовательно, расслабление представляет собой прекращение производства мышечного напряжения. Когда поперечные мостики отделяются во время расслабления, внутренняя эластичная сила, накопившаяся в филаментах во время сокращения, выделяется. Таким образом, эластическая тяга эластичных компонентов является именно тем, что возвращает длину миофибрилл к длине, наблюдаемой в состоянии несокращения (Gowitzke и Milner, 1988). Вторая возможная восстанавливающая сила может возникать в результате взаимного наложения тонких филаментов, которые отталкивают друг друга ввиду одинакового заряда. Предполагают, что «такие восстанавливающие силы снижают энергетическую стоимость расслабления» (Pollack, 1990).
38