

belyuchenko_i_s_smagin_a_v_i_dr_analiz_dannykh_i_matematiche
.pdf7. Проверка статистических гипотез. Когда выборки представлены несколькими повторностями, для проверки значимости различий между сообществами необходимо использовать дисперсионный анализ. Если непосредственно сравниваются результаты двух исследований, важно использовать один и тот же индекс разнообразия. По этой причине более информативным может оказаться использование индекса Шеннона. Достоверность различия коэффициентов разнообразия определяется по t-критерию Стьюдента.
Соответствие каждой теоретической модели рангового распределения реальным данным оценивается по двум критериям, взаимно дополняющим друг друга.
1. Коэффициент детерминации (квадрат коэффициента линейной корреляции) между реальными и теоретическими значениями обилия ( % общей дисперсии, объясненный моделью). Чем ближе его величина к 100 %, тем лучше соответствие модели.
2. Сумма квадратов отклонений реальных значений от модельных. Чем меньше эта величина, тем лучше соответствие модели. В случае, когда качество аппроксимации двумя (или более) моделями приблизительно одинаково по обоим указанным критериям, можно использовать любую из них.
Оценка параметров моделей и их ошибок с помощью компьютера. Модель «разломанного стержня» и логнормальное распределение являются беспараметрическими. Для оценки параметров модели геометрического распределения при расчетах без применения персонального компьютера используется процедура приведения их к линейному виду:
lg{n(k)} = lg(n1)+(k-1) lg(Ck), |
(11.31) |
где n(k) – относительное обилие вида с рангом k; n1, Ck, – параметры.
161
После линеаризации оценка параметров и их стандартных ошибок проводится методом наименьших квадратов для линейной регрессии. Необходимо различать ошибки параметров данной линии регрессии и ошибки, связанные с выборочными флуктуациями (различия между разными пробами, ограниченный объем выборок, погрешности в определении численностей и пр.). Для учета таких ошибок рекомендуется обрабатывать данные в нескольких параллельных пробах. При этом погрешности в параметрах ранговых распределений обычно бывают меньше, чем погрешность иных характеристик сообщества (таких, например, как общая численность, биомасса или число видов).
162
Глава 12. ДИНАМИЧЕСКИЕ ПОПУЛЯЦИОННЫЕ МОДЕЛИ
12.1Количественная оценка рождаемости, смертности, скорости роста популяции
Описание изменения численности популяции во времени составляет предмет популяционной динамики. При изучении динамики численности особей в популяциях, а также процессов, определяющих ее изменение, можно установить следующее выражение:
Nt = N0+В–D+С–Е, |
(12.1) |
где Nt – число особей в популяции в момент времени t;
N0 – число особей, находившихся на этой территории прежде; В – число особей, родившихся за время t;
D – число особей, погибших за то же время;
С и Е – число эмигрантов и иммигрантов за то же время.
Эти процессы в существовании любых популяций определяют основную задачу биоэкологии: описать, истолковать и понять закономерности распространения и динамику численности живых существ.
Зная динамические показатели популяции, можно судить о многих ее особенностях, условиях развития и, косвенно, о состоянии экосистемы в целом. Основные из них:
рождаемость, смертность, скорость роста популяции.
Скорость рождаемости, смертности, скорость роста по-
пуляции могут быть средними и мгновенными. Рождаемость (скорость рождения) – число новых
особей появившихся в популяции за единицу времени в результате размножения.
Различают максимальную, фактическую, абсолютную и удельную рождаемость.
163
Максимальная рождаемость – реализация возможности рождения при отсутствии лимитирующих факторов среды.
Фактическая рождаемость – реальная реализация воз-
можности рождения.
Абсолютная рождаемость (скорость рождаемости),
выражают отношением: dNn/dt, где dNn – число особей, родившихся за некоторый промежуток времени dt.
Удельная рождаемость (b) – отношение скорости рождаемости к исходной численности(N): b = dNn/(Ndt).
Смертность (скорость смертности) – число особей,
погибших в популяции за единицу времени (от хищников, болезней, старости и других причин).
Различают минимальную, фактическую, абсолютную и удельную смертность.
Минимальная смертность – минимально возможная величина смертности.
Фактическая смертность – реальная величина смертно-
сти.
Абсолютная смертность (скорость смертности) выражают отношением: dNm/dt, где dNm – число особей, погибших за промежуток времени dt.
Удельная смертность – (d) – отношение скорости смертности к исходной численности(N): d = dNm/(Ndt).
Скорость роста популяции – изменение численности популяции в единицу времени. Она зависит от показателей рождаемости, смертности и миграции (вселения – иммиграции и выселения – эмиграции) и может быть положительной, отрицательной и нулевой. Общая скорость роста популяции в отсутствие лимитирующего влияния среды зависит от возрастного состава и вклада в репродукцию различных возрастных групп.
Различают абсолютную и удельную скорости роста.
164
Абсолютная (общая) скорость роста выражается от-
ношением: dN/dt, где dN – изменение численности популяции за промежуток времени dt.
12.2 Биотический потенциал популяции
Показатель наследственно обусловленной способности популяции к росту и называется биотическим потенциалом (репродуктивным потенциалом) вида:
r |
dN |
|
d ln( N ) |
|
или |
dN |
rN |
(12.2) |
|
Ndt |
dt |
dt |
|||||||
|
|
|
|
|
|||||
Величина биотического потенциала различна у разных видов. Например, самка косули способна производить на свет 10 – 15 козлят, самка медоносной пчелы – 50 тыс. яиц, рыба-луна – до 3 млрд. икринок.
В природе в связи с действием лимитирующих факторов биотический потенциал популяции никогда не реализуется полностью. Его величина обычно складывается как разность между рождаемостью и смертностью в популяции:
r = b– d, |
(12.3) |
где b – число родившихся;
d – число погибших особей в популяции за один и тот же период времени.
Когда b = d, r = 0 и популяция находится в стационарном состоянии. Когда b > d, r >0, численность популяции увеличивается. Когда b < d, r < 0, численность популяции сокращается. Формула d = b – r позволяет определить смертность, которую трудно измерить непосредственно, а определить r достаточно просто непосредственными наблюдениями.
165
Пример. Популяция состоит из 800 особей. В среднем за год рождается 150 особей, а умирает 50. Определить скорость естественного увеличения популяции (r).
Решение: r = 150/800 – 50/800 = 0,125 /год.
12.3 Продолжительность жизни и выживаемость
Длительность существования особи называется продолжительностью жизни. Различают физиологическую, максимальную и среднюю продолжительность жизни.
Физиологическая продолжительность (ФПЖ) – это продолжительность жизни, без влияния на особей лимитирующих факторов. Она возможна только теоретически.
Максимальная продолжительность жизни (МПЖ) –
это продолжительность жизни в реальных условиях среды. Она варьирует от нескольких минут у бактерий до нескольких тысячелетий у древесных растений (секвойя).
Средняя продолжительность жизни (СПЖ) – это сред-
нее арифметическое продолжительности жизни всех особей. Выживаемость – абсолютное число особей (или процент от исходного числа особей), сохранившихся в популя-
ции за определенный промежуток времени:
Z |
n |
100 % , |
(12.4) |
N |
|
||
|
|
|
где Z – выживаемость, %; n – число выживших;
N – исходная численность популяции.
Выживаемость зависит от ряда причин: возрастного и полового состав популяции, действия тех или иных факторов среды и др.
166
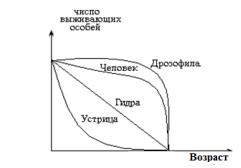
12.4Демографические таблицы популяций, кривые выживания
Один из методов статистического анализа популяций – построение демографических таблиц (таблиц выживания), в которых приводится возрастная структура по продолжительности жизни особей. Таблицы выживания бывают динамические и статические.
Динамические таблицы строятся по данным прямых наблюдений за жизнью когорты с помощью регистрации возраста наступления смерти всех членов данной когорты. Такие таблицы требуют длительного наблюдения, измеряемого месяцами или годами. Практически невозможно такую таблицу сделать для долго живущих животных или для человека – для этого понадобится более 100 лет.
Статические таблицы выживания составляются по данным наблюдений за относительно короткий промежуток времени за смертностью в отдельных возрастных группах. Зная численность этих групп можно рассчитать смертность, специфическую для каждого возраста.
На основании анализа демографических таблиц можно строить кривые выживаемости особей в популяции, которые устанавливают зависимость количества выживших особей от их возраста.
Рисунок 12.1 – Кривые выживаемости
167
Выделяют три типа кривых выживаемости (рисунок
12.1):
–сильновыпуклые кривые – соответствуют ситуации, когда смертность ничтожно мала в молодом и среднем возрасте, но быстро увеличивается в старом возрасте, и все особи погибают за короткий срок. Пирл назвал эту кривую «кривой дрозофилы». Такой тип кривых выживания характерен для многих видов крупных животных, в том числе и для человека;
–кривые диагонального типа – относятся к тем видам, у
которых смертность во всех возрастных группах одинаковая. Такова динамика многих рыб, пресмыкающихся, птиц, многолетних травянистых растений.
–сильновогнутые кривые – характерны для видов,
смертность которых чрезвычайно высока на ранних стадиях развития, а для выживших особей становится низкой. Эту кривую Пирл назвал «типом устрицы». Ей соответствует возрастная динамика большинства видов деревьев. Например, семена дуба при прорастании погибают в больших количествах, а гибель укоренившихся растений остается чрезвычайно низкой.
12.5 Оценка скорости роста популяции
Чтобы рассчитать скорость роста популяции, необходимо знать величину так называемой чистой скорости вос-
производства (R0) и среднее время генерации.
Чистая скорость воспроизводства показывает, во сколько раз увеличится численность популяции за одно поколение (или ожидаемое число потомков женского рода, которых родит данная новорожденная самка за всю свою жизнь). В таблице 12.1 ожидаемое число потомков рассчи-
тывается как произведение выживаемости на плодовитость. Сумма ожидаемого числа потомков и есть чистая скорость
168
размножения R0. В рассматриваемом примере R0 = 3,1. Это означает, что к моменту гибели самки в популяции произведут в среднем 3,1 потомка женского пола.
Таблица 12.1 – Расчет чистой скорости размножения и среднего времени генерации
|
Выжи- |
Плодовито |
Ожидаемое |
Произведение воз- |
|
|
число по- |
раста на ожидаемое |
|||
Возраст, t |
ваемость |
сть |
|||
томков |
число потомков |
||||
|
P(t) |
N(t) |
|||
|
P(t)· N(t) |
t· P(t)· N(t) |
|||
|
|
|
|||
0 |
1,00 |
0,0 |
0,0 |
0,0 |
|
1 |
0,50 |
1,0 |
0,5 |
0,5 |
|
2 |
0,40 |
3,0 |
1,2 |
2,4 |
|
3 |
0,30 |
4,0 |
1,2 |
2,4 |
|
4 |
0,10 |
2,0 |
0,2 |
0,8 |
|
5 |
0,00 |
0,0 |
0,0 |
0,0 |
|
Чистая скорость размножения R0 |
3,1 |
- |
|||
Общий взвешенный возраст |
- |
6,1 |
|||
Среднее время генерации – время жизни популяции (или средний возраст, в котором самки производят потомство). Чем раньше появятся детеныши, тем раньше они, в свою очередь произведут потомство, тем выше будет идти рост популяции.
Среднее время генерации T вычисляют, находя средний возраст, в котором самка рожает детенышей. Для этого необходимо общий взвешенный возраст разделить на чистую скорость размножения. В рассматриваемом примере имеем:
Т = 6,1/3,1 = 1,93. Следовательно, популяция будет увеличиваться в 3,1 раза каждый 1,93 года.
Скорость роста популяции оценивается по формуле:
r |
ln( R0 ) |
. |
(12.5) |
|
|
||
|
T |
|
|
В нашем примере: r = ln3,1/1,93 = |
0,5862. |
||
Размерность для r –«единица времени»-1.
169
Легко найти r, зная время удвоения популяции. В этом
случае: |
r ln 2 /T . |
|
|
|
Полезно помнить формулы, связывающие T, R0 и r: |
||||
R |
erT , отсюда |
T |
ln( R0 ) |
. |
0 |
|
r |
||
|
|
|
||
Для оценки r на практике очень часто приходится пользоваться формулой:
r |
ln Nt |
|
ln Nt |
, |
|
|
2 |
1 |
(12.6) |
||
t |
|
t |
|||
|
|
|
|
||
|
|
2 |
1 |
|
|
где Nt2 – численность популяции в момент t2,
Nt1 – численность в момент t1.
Например, если за 2 часа численность популяции изменилась с Nt1 = 10 до Nt2 = 100, то
r |
ln100 ln10 |
|
4,6 2,3 |
1,15 час-1. |
|
|
|
|
|||
2 |
|
2 |
|
|
|
12.6Модель экспоненциального роста популяции Мальтуса
Принцип экспоненциального роста численности популяции (Свирежев, Логофет, 1978) – один из основных экологических принципов динамики популяций. Его используют в экологии, прежде всего, для того, чтобы количественно охарактеризовать потенциальные возможности популяции к росту и выявить факторы, ограничивающие рост изучаемой популяции.
Модель экспоненциального роста предложена Мальтусом в 1798 г. в его классическом труде «О законе роста народонаселения». Томас Роберт Мальтус (1766–1834) – из-
170