

40.Переваривание и всасывание липидов .
В ротовой полости липиды практически не перевариваются , но язычными железами выделяется липаза , которая в течении 2 – 4 часов расщепляет 20 – 25 % липидов .
Желудочная липаза тоже малоактивна потому , что оптимум рН для неё 5 – 7,5 , а в желудке – 1,5 . Кроме этого липаза активно гидролизует предварительно эмульгированные жиры , а в желудке условий для эмульгирования нет .
Переваривание жиров в полости желудка играет важную роль у детей , особенно у грудных . У них рН желудочного сока = 5,0 . Это способствует перевариванию эмульгированного жира молока желудочной липазой . Кроме того у детей грудного возраста наблюдается адаптивное усиление синтеза желудочной липазы . Здесь же продукты гидролиза липидов всасываются в стенку желудка и по портальной вене идут в печень . Расщепление жиров у взрослого человека происходит в проксимальном отделе тонкого кишечника .В двенадцатиперстной кишке прежде всего происходит нейтрализация соляной кислоты желудочного сока бикарбонатами панкреатического и кишечного соков . Выделяющиеся при разложении бикарбонатов пузырьки углекислого газа способствуют хорошему перемешиванию химуса . Одновременно начинается эмульгирование . Дело в том . что липиды в большинстве своём гидрофобны , тогда как внутренняя среда организма гидрофильна , поэтому нужно придать липидам гидрофильную форму путём эмульирования . сущность эмульгирования – снижение поверхностного натяжения и распад жиров на мелкие сферы . Главный фактор эмульгировния – желчные кислоты и их соли , которые являются производными холановой кислоты . Основная масса желчных кислот находится в коьюгированном виде : связаны либо с глицином , либо с таурином . Синтез желчных кислот из холестерина происходит в цитоплазме гепатоцитов путём микросомального гидроксилирования . В желчи человека содержится холевая кислота ( наибольшее количество ) , дезоксихолевая , хенодезоксихолевая кислоты .

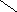





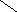

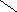
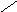
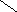
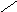

 СН3
СН3


 СН3 |
СН3 |
СН – СН2 – СН2 – СООН



 |
|
СН3
Холановая кислота
СН3
|

 ОН
СН – СН2
– СН2 – СООН
ОН
СН – СН2
– СН2 – СООН

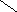


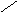
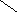
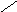


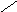
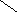

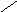


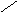
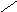



 СН3
СН3


 СН3
СН3
НО ОН

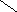


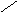

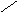
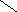

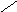
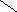

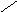




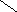



 Холевая
кислота
Холевая
кислота
СН3
|
ОН СН – СН2 – СН2 – СООН
СН3
СН3
НО
Дезоксихолевая кислота
 CН3
– СН – СН2 –
СООН
CН3
– СН – СН2 –
СООН

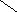
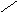

СН3



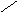
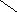




 СН3
СН3




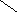
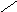
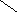


НО НО
Хенодезоксихолевая кислота .
Считается , что только комбинация : соль желчной кислоты + ненасыщенная жирная кислота + моноглицерид способна дать необходимую степень эмульгирования
Функции желчных кислот :
1) обеспечивают эмульгирование жира , а также стабилизируют уже образовавшуюся эмульсию .
2) сменяют желудочное пищеварение на кишечное , то есть инактивируют пепсин .
3) подавляют гнилостные процессы в кишечнике .
4) активируют панкреатическую липазу .
5) транспортируют жирные кислоты через биологические мембраны энтероцитов .
6) усиливают секреторно-моторную деятельность кишечника .
Количество выделяемых желчных кислот в 5 раз ниже требуемого , однако дефицита никогда нет так как существует печёночно-кишечный круговорот компонентов желчи ( желчных кислот прежде всего ). За сутки совершается 5 оборотов : желчные кислоты (90-95%) реабсорбируются в тонком кишечнике и через портальную вену поступают обратно в печень .То есть этот оборот облегчает работу печени по синтезу компонентов желчи , способствуя тем самым выполнению ею других функций ( обменных , защитных ).
Основной фермент , осуществляющий гидролиз жиров , - панкреатическая амилаза , активируемая желчными кислотами . Активирующее влияние этих кислот на липазу выражается в смещении оптимума , действие данного фермента с рН 8,0 до 6,0 , то есть до той величины рН ,которая поддерживается в двенадцатиперстной кишке . Предполагается также , что активация пролипазы идёт путём образования комплекса с колипазой ( кофактор) в молярном соотношении 2:1. Это и способствует сдвигу рН с 8,0 до 6,0 . Путь же активации – частичный протеолиз .
Есть основания считать , что существует липаза 2-х типов , одна специфична в отношении связей в положениях 1 и 3 , другая в положении 2 .
Гидролиз триглицеридов идёт постадийно : сначала быстро гидролизуются связи 1 и 3 , затем медленно идёт гидролиз 2-моноглицерида .
O
||
O CH2 – O – C – R1
|| |
R2 – C – O – CH O
| ||
CH2 – O – C – R3


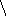 H2O
H2O
H2O
H2O


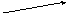 R1COOH
R3COOH
R1COOH
R3COOH
O
||
O CH2OH O CH2 – O – C – R1
|| | || |
R2 – C – O – CH O R2 – C – O – CH
| || |
 CH2
– O – C – R3
CH2OH
CH2
– O – C – R3
CH2OH
Диглецирид диглецирид

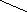


 H2O
H2O
H2O
H2O
R3COOH R1COOH
CH2OH O
| ||
CH – O – C – R2
|
 CH2OH
CH2OH
2-моноглицерид
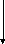
 H2O
H2O
R2COOH
CH2OH
|
CHOH
|
CH2OH
глицерин
В расщеплении жиров принимает участие и кишечная липаза , однако активность её мала и к тому же она гидролизует расщепление только моноглицеридов . То есть продукты гидролиза триглицеридов : жирные кислоты , моноглицериды и глицерин .
Всасывание.
Только эмульгированные жиры частично всасываются без предварительного гидролиза .
Продукты гидролиза всасываются так :
-жирные кислоты с короткой углеродной цепью (менее С12) и глицерин будучи хорош растворимы в воде , свободно всасываются в стенку кишечника , а затем через портальную вену в печень .
-жирные кислоты длинной цепью и моноглицериды всасываются с помощью желчных кислот ,фосфолипидами и холестерином мицеллы . Структура этих мицелл такова , что их гидрофобное ядро ( жирные кислоты и моноглицериды ) оказывается окружёнными гидрофильной оболочкой из желчных кислот и фосфолипидов . Предполагается , что всасывание мицелл происходит путём мицеллярной диффузии или путём пиноцитоза .
Переваривание и всасывание фосфолипидов и холестерина .
Расщепление фосфолипидов осуществляется под действием фосфолипаз панкреатического сока . Различают несколько типов фосфолипаз .
Фосфолипаза А1 гидролизует эфирную связь в положении 1 , в результате чего отщепляется 1 молекула жирной кислоты .
Фосфолипаза А2 гидрлизует отщепление жирных кислот в положении 2 .
Образующиеся при этом продукты называются лизофосфолипидами , в частности при гидролизе фосфатидилхолина образуется лизофосфатидилхолин .Они токсичны и вызывают разрушение мембран клеток . именно поэтому яд змей , содержащий высокоактивную фосфолипазу А2 , гемолизирует эритроциты . Однако токсичность лизофосфолипидов в кишечнике устраняется одновременным действием на глицероффосфолипиды и фосфолипазы А1 и А2 . В результате образуется нетоксичный продукт .
Фосфолипаза С вызывает гидролиз связи между остатками глицерина и Н3РО4 .
Фосфолипаза Д расщепляет эфирную связь между азотистым основанием и Н3РО4 . В результате образуется азотистое основание и Н3РО4 .
O

 || Ф
А1
R1COOH
|| Ф
А1
R1COOH
CH2 – O – C – R1

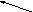 O
O
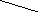
 ||
||
 СH – O
– C – R2
Ф А2
R2COOH
ГЛИЦЕРИН
СH – O
– C – R2
Ф А2
R2COOH
ГЛИЦЕРИН
 O
O
 || Ф С
|| Ф С
 СH2
– O – P – O
– CH2
– CH2
– N(CH3)3
H3PO4
СH2
– O – P – O
– CH2
– CH2
– N(CH3)3
H3PO4

 |
|
OH Ф Д
фосфатидилхолин холин
Н3РО4 всасывается в виде Na или К солей .Азотистые основания ( холин ) всасывается в виде своих активных форм .
Что касается холестерина , то он попадает в ЖКТ с яичным желтком , мясом , печенью . холестерин поступает в количестве 0,1 – 0,3 г в свободном виде или в форме эфиров . Эфиры расщепляются на холестерин и жирные кислоты при участии фермента панкреатического и кишечного сока – холестеролэстеразы . холестерин , нерастворимый в воде , всасывается только в присутствии желчных кислот .