

39. Липиды . Строение , классификация , биологическая роль .
Липиды – большая группа веществ , разнородных по составу , но объеденённых в одну группу по трём признакам :
-
гидрофобность
-
растворимы в органических растворителях
-
метаболизм в организме
Классификация.
Выделяют две большие группы липидов по их отношению к гидролизу :
-
омыляемые
-
неомыляемые
Омыляемые липиды – производные жирных кислот .

 Простые
Сложные
Простые
Сложные
(

 воска , масла , жиры )
воска , масла , жиры )
гликолипиды

 R1
СН2 –
О – СОR1
R1
СН2 –
О – СОR1
| | фосфолипиды
 R
– O – C
СН – О – СОR2
цереброзиды ганглиозиды
R
– O – C
СН – О – СОR2
цереброзиды ганглиозиды
|| | сульфатиды
O СН – О – СОR3


 глицерофосфолипиды сфинголипиды
глицерофосфолипиды сфинголипиды

фосфотидилхолины плазмогены сфингомиелины церамиды
фосфотидилэтанол фосфотидилинозины
амины
фосфотидилсерины
Ненасыщенные липиды – производные изопрена .

Животного происхождения : Растительного происхождения :
Стероиды : каротиноиды ( а , в )
Стерины ( холестерин ) Vit A , E , K .
Стериды ( ненасыщенные жирные кислоты )
Краткое строение :
Все жирные кислоты можно разделить на три группы :
-насыщенные
-мононенасыщенные
-полиненасыщенные
В состав омыляемых липидов входят жирные кислоты с числом атомов от 4 до 28 ( чаще 16 - 20 ), имеют неразветвлённую углеродную цепь и чётное число атомов .
Насыщенные жирные кислоты :
-пальмитиновая ( С15Н31СООН )
-стеариновая ( С17Н35СООН )
-арахидоновая ( С19Н39СООН )
Мононенасыщенные ( с одной двойной связью )
-олеиновая ( С17Н33СООН )
-кротоновая ( С3Н5СООН )
-пальмитиновая ( С15Н29СООН )
Полиненасыщенные:
-линолевая ( С17Н31СООН ) 2 двойные связи
-линоленовая ( С17Н29СООН) 3 двойные связи
-арахидоновая ( С19Н31СООН ) 4 двойные связи
Все ненасыщенные жирные кислоты находятся в цис – положении и поэтому биологически активны . Организм синтезирует насыщенные жирные кислоты с 1двойной связью , с двумя и более связями – поступают с пищей .
Воска – сложные эфиры высших жирных кислот и одноатомных спиртов . Могут входить в состав жира , покрывающего кожу . Нейтральные жиры ( глицериды ) – эфиры глицерина и жирных кислот : если жирными кислотами этерефицируются все три гидроксильные группы глицерина , то это триглицерид , если два – то диглицерид , если один – то моноглицерид . Нейтральные жиры находятся в организме либо в форме протоплазматического жира , либо в форме резервного жира . Количество первого строго определено и не изменяется даже при патологическом ожирении ; количество резервного жира подвергается колебаниям . Различают простые и смешанные триглицериды . Простые содержат остатки одинаковых жирных кислот , смешанные – разных жирных кислот .
Фосфолипиды .
Глицерофосфолипиды – производные фосфатидной кислоты , общая формула :
О
||
О CН2 – О – С – R
|| |
R2 – С – О – СН О
| ||
СН2 – О – Р – О – Х
|
ОН
Е сли
Х = СН2 – CН2
– NH2
фосфотидилэтаноламины
( кефалины )
сли
Х = СН2 – CН2
– NH2
фосфотидилэтаноламины
( кефалины )
Е сли
Х = СН2 – СН2N(
СН3 ) 3
фосфотидилхолины
сли
Х = СН2 – СН2N(
СН3 ) 3
фосфотидилхолины
Е сли
Х = СН2 – СН –
NH2
фосфотидилсерины
сли
Х = СН2 – СН –
NH2
фосфотидилсерины
|
СООН
Плазмогены содержатся в больших количествах в мышцах и эритроцитах .
О CН – О = СН = СН – R1
|| |
R – С – О – СН О
| ||
СН2 – О – Р – ( холин или коламин )
|
ОН
Фосфатидилинозитолы – входят в состав мозга , печени ,лёгких.
О
||
СН2 – О – С – R
|
R2 – C – О – CН О
| ||
 CН2
– О – Р – О
CН2
– О – Р – О
| ОН ОН
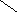


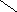
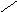 ОН |
|
ОН |
|
ОН
| |
| ОН
ОН
Фосфатидная кислота О
||
О CН2 – О – С – R1
|| |
R2 – С – О – СН О
| ||
СН2 – О – Р – Н
|
ОН
Сфинголипиды .
Сфингомиелины - входят в состав мембран нервной ткани , печени , почек . Имеют в составе сфингозин :
СН3 – ( СН2 ) 12 – СН = СН – СН – СН – СН2ОН
| |
ОН NH2
Церамид : СН3 – (СН2 )12 – СН = СН – СН – СН – СН2ОН
| |
 ОН
NHСОR1
остаток жирной кислоты
ОН
NHСОR1
остаток жирной кислоты
Гликолипиды .
Цереброзиды СН3 – ( СН2 ) 12 – СН = СН – СН – СН – СН2 – О
 | |
| |
ОН NH
|
CO
|
R
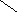

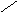
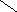

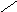
 СН2ОН
СН2ОН
НО О

 ОН
ОН

ОН
Ганглиозиды :
Гематозид ( строма эритроцитов ) :


 сфингозин глюкоза
галактоза
N –
ацетилнейраминовая кислота
сфингозин глюкоза
галактоза
N –
ацетилнейраминовая кислота
Стероиды .
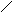












 Общая структура
Общая структура
СН3

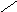
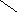
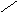
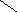

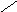 СН3
СН3
СН3

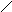


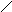

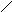


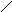


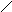



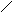



 Холестерин :
СН3
Холестерин :
СН3

 СН3
СН3

НО
Функции липидов :
-
энергетическая ( 1 г – 39 кДж )
-
незаменимый фактор пмтания ( Vit A , D , E , K )
-
защитная ( теплоизоляция , защитная )
-
структурная
-
формообразующая
-
и
 сточник
воды ( при окислении 100 г жира
107 г воды )
сточник
воды ( при окислении 100 г жира
107 г воды )
-
растворитель для Vit A , D , E , K .
-
смазка для кожи
-
регуляторная ( липиды принимают участие в регуляции активности ферментов , процессов транспорта , осуществляют контроль за реакциями биологического окисления и энергетического обмена в клетке , обеспечивают компартментализацию обмена в клетке , участвуют в межмолекулярных взаимодействиях и обеспечении молекулярных механизмов памяти )
-
модуляторы белков и ферментов
-
влияют на проницаемость мембран , участвуют в передаче нервного импульса, создании межклеточных контактов .
Строение мембран .
Молекула фосфолипидов амфолитна : одна её часть гидрофобная ( радикалы R1 и R2 ), другая часть гидрофильна , благодаря « – » заряду остатка фосфорной кислоты и « + » заряду радикала R3 .То есть фосфолипиды обладают полярными свойствами , поэтому при смешивании в водной фазе они способны к самосборке и образованию бислоя в котором гидрофобные части взаимодействуют между собой , а гидрофильные с водой так как существование монослоя термодинамически не выгодно . В состав фосфолипидов входят ненасыщенные жирные кислоты , которые придают им , при температуре тела 0 жидковатую консистенцию ( консистенция всех мембран приближается к консистенции сливочного масла ) В состав бислоя входят молекулы холестерина ( в гидрофильной части ) . Соотношение холестерина и фосфолипидов должно быть 1 : 1 , если холестерина больше , то увеличивается вязкость бислоя и мембрана утрачивает свои свойства .
Первая модель мембраны – бутербродная , была предложена в 1925 году . В зависимости от взаимного расположения фосфолипидов и белков существует много моделей мембран :
1) мозаичная – белки расположены либо внутри , либо на поверхности бислоя ( внутренняя митохондриальная мембрана )
2) ковровая – мембрана на половину состоит из липидов . Белки насквозь пронизывают толщу мембраны , образуя поры , каналы . В гидрофобную фазу мембраны погружены гидрофобные части белков ( валин , лейцин , изолейцин )
-
глобулярная
мембраны имеют неоднородный фосфолипидный состав . Внутренний и внешний слои мембраны могут быть образованы различными фосфолипидами . Кроме того молекулы белка с гидрофобными концами делают мембрану похожей на фильтр , который поддерживают определённую концентрацию ионов и клеточный метаболизм всецело зависит от функционировании этого фильтра .