

Гормоны и их физиологические эффекты
В этом разделе дана физиологическая характеристика различных гормонов, синтезируемых и секретируемых клетками эндокринной системы.
Гипоталамо-гипофизарная система
Часть промежуточного мозга — гипоталамус — и отходящий от его основания гипофиз анатомически и функционально составляют единое целое — гипоталамо-гипофизарную эндокринную систему (рис. 18–1, 18–2, 18–3, см. также рис. 16–3,В,Г).
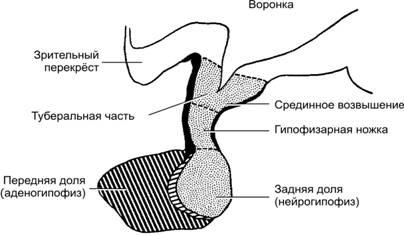
Рис.18–1.Анатомическиевзаимоотношениягипоталамусаиножкигипофиза[11].Точечнаяштриховка: срединное возвышение и задняя доля гипофиза (нейрогипофиз) имеют нейральное происхождение и фактически являются частью гипоталамуса.Косаяштриховка: эпителиальная часть гипофиза (аденогипофиз) развивается из эктодермы ротовой бухты. Роль гипоталамо–гипофизарной системы для эндокринной регуляции функций организма столь велика, что её иногда называют «президентом эндокринного общества».
Гипоталамус
В нейросекреторных нейронах гипоталамуса синтезируются нейропептиды, поступающие как в переднюю (рилизинг–гормоны), так и в заднюю (окситоцин и вазопрессин) доли гипофиза.
Рилизинг–гормоны
Гипоталамические рилизинг–гормоны (от англ. releasinghormone) — группа нейрогормонов, мишенями которых являются эндокринные клетки передней доли гипофиза. С функциональной точки зрения рилизинг–гормоны подразделяют на либерины (рилизинг–гормоны, способствующие усилению синтеза и секреции соответствующего гормона в эндокринных клетках передней доли гипофиза) и статины (рилизинг гормоны, подавляющие синтез и секрецию гормонов в клетках–мишенях). К гипоталамическим либеринам относятся соматолиберин, гонадолиберин, тиреолиберин и кортиколиберин, а статины представлены соматостатином и пролактиностатином.
Соматостатин— циклический тетрадекапептид C76H104N18O19S2(мол. масса 1637,9) — синтезируется многими нейронами ЦНС (в том числе нейронами околожелудочковой области гипоталамуса),‑клетками островковЛангерхансаподжелудочной железы, эндокринными клетками ЖКТ и ряда других внутренних органов.
Функции. Соматостатин — мощный регулятор функций эндокринной и нервной систем,ингибируетсинтез и секрецию множества гормонов и секретов.
Передняядолягипофиза. Соматостатин подавляет синтез и секрецию гормона роста, АКТГ и тиреотропного гормона.
ОстровкиЛангерханса. Соматостатин подавляет секрецию инсулина иглюкагона.
Энтероэндокриннаясистема. Соматостатин ингибирует секрецию гастрина, холецистокинина, секретина.
Почка. Соматостатин подавляет секрецию ренина.
Желудок. Соматостатин ингибирует желудочную секрецию.
Рецепторы. Соматостатин реализует эффекты через связанные с G‑белком мембранные высокоаффинные рецепторы (ингибированиеаденилатциклазыцАМФ ...уменьшение [Ca2+] в цитозолеблокада экзоцитоза секреторных пузырьков). Идентифицировано не менее 5 подтипов рецепторов соматостатина, имеющих выраженную органную специфичность распределения.
Соматолиберин(соматокринин) — пептид, содержащий 44 аминокислотных остатка, синтезируется нейросекреторными нейронами дугового ядра (n.arcuatus) гипоталамуса и некоторыми опухолями островковых клеток поджелудочной железы (соматолибриномы).
Функция. Гипоталамический соматолиберин стимулирует секрецию гормона роста в передней доле гипофиза.
Рецепторсоматолиберина относят к семейству связанных с G‑белком рецепторов типасекретина(стимуляцияаденилатциклазыцАМФпротеинкиназа Aактивация кальциевых каналов типа Lувеличение [Ca2+] в цитозолеэкзоцитоз секреторных пузырьков).
Гонадолиберин(люлиберин) ипролактиностатин. ГенLHRHкодирует аминокислотные последовательности для гонадолиберина и пролактиностатина.
Мишенигонадолиберина — гонадотрофы, а пролактиностатина — лактотрофы передней доли гипофиза.
Функции. Гонадолиберин — ключевой нейрорегулятор репродуктивной функции,стимулируетсинтез и секрецию ФСГ и ЛГ в продуцирующих гонадотрофы клетках, а пролактиностатинподавляетсекрецию пролактина из лактотрофных клеток передней доли гипофиза.
Рецепторылюлиберина — трансмембранные гликопротеины, связанные с G‑белком. Активирование G‑белка приводит к образованию инозитолтрифосфата и диацилглицерола, что в конечном итоге стимулирует секрецию ФСГ и ЛГ.
Тиреолиберин— трипептид, синтезируется многими нейронами ЦНС (в том числе нейросекреторными нейронами паравентрикулярного ядра).
Мишенитиреолиберина — тиреотрофы и лактотрофы передней доли гипофиза.
Пролактин. Тиреолиберинстимулируетсекрецию пролактина из лактотрофов.
Тиреотропный гормон. Тиреолиберинстимулируетсекрецию тиреотропина из тиреотрофов.
СТГ. В патологических ситуациях тиреолиберинстимулируетсекрецию гормона роста из соматотрофов.
Рецепторы. Связывание тиреолиберина с рецепторами активирует в клетках–мишенях G‑белок, что приводит к активации фосфолипазы A2и синтезу производных арахидоновой кислоты.
Кортиколиберин— пептид из 41 аминокислотного остатка — синтезируется в нейросекреторных нейронах паравентрикулярного ядра гипоталамуса, плаценте, Т-лимфоцитах.
Влияниеглюкокортикоидовнасинтезкортиколиберина
Гипоталамус. Глюкокортикоидыподавляютсинтез гипоталамического кортиколиберина.
Плацента. Глюкокортикоидыстимулируютсинтез плацентарного кортиколиберина.
Функциикортиколиберина многочисленны.
Передняядолягипофиза. Стимуляция синтеза и секреции АКТГ и других продуктов экспрессии гена проопиомеланокортина.
Стресс. Кортиколиберин — координатор эндокринных, нейровегетативных и поведенческих ответов в стрессовых ситуациях. Последние всегда затрагивают ось «кора головного мозга–гипоталамус–гипофиз–надпочечник».
Плацента. На сроке 18 нед беременности по содержанию в крови кортиколиберина можно выделить группы беременных с преждевременными и поздними родами. К предсказанному сроку содержание в крови кортиколиберина резко возрастает. Возможно, этот гормон является сигналом начала родов.
Рецепторыкортиколиберина относят к семейству связанных с G‑белком рецепторов типасекретина(стимуляция аденилатциклазыцАМФпротеинкиназа Aактивация кальциевых каналов типа Lувеличение [Ca2+] в цитозолеэкзоцитоз секреторных пузырьков).
Меланостатин—L-пролил-L-лейцилглицинамид — подавляет образование меланотропинов.
Либерины и статины по аксонам гипоталамических нейронов достигают срединного возвышения, где секретируются в кровеносные сосуды портальной системы кровотока (рис. 18–2), далее по воротным венам гипофиза эти нейрогормоны поступают в переднюю долю гипофиза и регулируют активность её эндокринных клеток (табл. 18-1, см. также рис. 18–4).
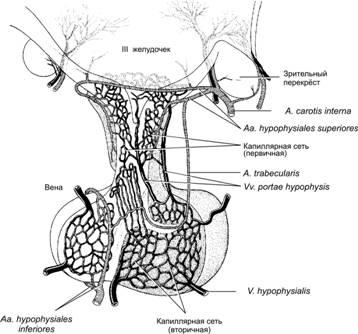
Рис.18–2.Гипоталамическаярегуляцияаденогипофиза[11]. Рилизинг–гормоны по аксонам нейросекреторных нейронов поступают в срединное возвышении, где секретируются в капилляры первичной капиллярной сети. Эти капилляры объединяются в несколько воротных вен гипофиза (vv.portaehypophysis), которые в передней доле гипофиза образуют вторичную капиллярную сеть.
Таблица18–1.Эффектыгипоталамическихнейрогормоновнасекрециюгормоноваденогипофиза
|
Нейрогормон |
Гормонгипофиза |
Эффект |
Примечание |
|
ТТГ-РГ |
ТТГ, пролактин |
|
трипептид |
|
Кортиколиберин |
АКТГ |
|
пп, 41 АК |
|
Соматолиберин |
СТГ |
|
пп, 44 АК |
|
Пролактиностатин |
Пролактин |
|
пептид |
|
Меланостатин |
Меланотропины |
|
амид трипептида |
|
Соматостатин |
СТГ, ТТГ, АКТГ |
|
п, 14 АК |
|
Люлиберин |
Лютропин (ЛГ), ФСГ |
|
декапептид |
Сокращения: АК — аминокислота, пп — полипептид, п — пептид.
Рольдофамина. Промежуточный продукт метаболизма тирозина и предшественник норадреналина и адреналина — катехоловый амин дофамин (3-гидрокситирамин), поступающий к клеткам передней доли гипофиза через кровь — ингибирует секрецию ФСГ, лютропина (лютеинизирующий гормон), тиреотропного гормона (ТТГ) и пролактина.
Гормоны задней доли гипофиза
Нанопептиды вазопрессин и окситоцин синтезируются в перикарионах нейросекреторных нейронов паравентрикулярного и супраоптического ядер гипоталамуса, транспортируются по их аксонам в составе гипоталамо-гипофизарного пути в заднюю долю гипофиза, где и секретируются в кровь (рис. 18–3, см. также рис. 16–3Г). Сигналом к секреции является импульсная активность этих же самых нейросекреторных нейронов.
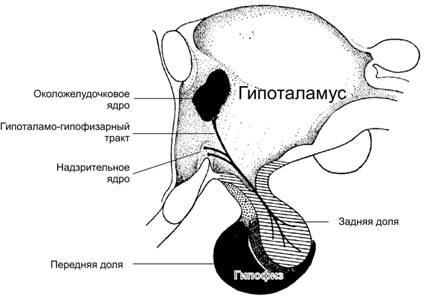
Рис.18–3.Гипоталамо-гипофизарныйтракт. Синтезируемые в телах нервных клеток супраоптического (nucleussupraopticus) и паравентрикулярного (n.paraventricularis) ядер гипоталамуса окситоцин и АДГ транспортируются по аксонам этих нейронов в заднюю долю гипофиза, где поступают в кровь.
Вазопрессин(aргинин вазопрессин, антидиуретический гормон — АДГ, C46H65N15O12S2) оказываетантидиуретический(регулятор реабсорбции воды в канальцах почки) исосудосуживающий(вазоконстриктор)эффекты(эти эффекты гормона приводят к повышению системного АД). Главная функция АДГ —регуляцияобменаводы(поддержание постоянного осмотического давления жидких сред организма), что происходит в тесной связи с обменом натрия.
Экспрессия гена АДГ происходит в части нейросекреторных нейронов паравентрикулярного и супраоптического ядер гипоталамуса. Внегипоталамическая секреция АДГ возможна в клетках злокачественных опухолей (например, овсяно–клеточная карцинома лёгкого, рак поджелудочной железы).
Секрецию АДГ стимулируют: гиповолемия через барорецепторы каротидной области, гиперосмоляльность через осморецепторы гипоталамуса, переход в вертикальное положение, стресс, состояние тревоги.
Секрецию АДГ подавляют: алкоголь,–адренергические агонисты, глюкокортикоиды.
Окситоцин. Экспрессия гена окситоцина и гена АДГ происходит в многоотростчатых нейронах супраоптического и паравентрикулярного ядер гипоталамуса, но в отдельных группах нервных клеток.
Регулятор секреции окситоцина и АДГ — импульсная активность аксонов нейросекреторных нейронов. При этом окситоцин, как и АДГ, отщепляется от нейрофизинов и поступает в кровь.
Окситоцин секретируется также при раздражении соска и околососкового поля молочной железы.
Мишени окситоцина — ГМК миометрия и миоэпителиальные клетки молочной железы.
Окситоцин стимулируетсокращение ГМК миометрия в родах, при оргазме, в менструальную фазу месячного цикла
Окситоцин также стимулируетсокращение миоэпителиальных клеток альвеол лактирующей молочной железы (рефлекс молокоотделения).
Рецептор окситоцина — трансмембранный гликопротеин, связанный с G‑белком. Взаимодействие окситоцина с его рецептором приводит к стимуляции фосфолипазы С, образованию фосфатидилинозитола и увеличению содержания внутриклеточного Ca2+.