

Справочный материал по Физиологии.
Глава 14 – Контрольпозыидвижений.
Мотонейроны спинного мозга и нейроны двигательных ядер черепных нервов — общий конечный путь к скелетным мышцам. Именно на этих мотонейронах конвергируют сигналы из множества источников. Нервные пути, конвергирующие к мотонейронам, выполняют три взаимосвязанных функции: сигнализируют о произвольной активностиподготавливают позу тела к устойчивому состоянию для движениякоординируют действия различных мышц для осуществления плавных и точных движений. Программа произвольных движений формируется в структурах мозга, а команды поступают к мышцам по пирамидному пути (корково–спинномозговые, корково-ядерные, корково-ретикулярные и другие нервные волокна). На рис. 14–1 показаны взаимодействия между основными структурами ЦНС в ходе осуществления произвольных движений.
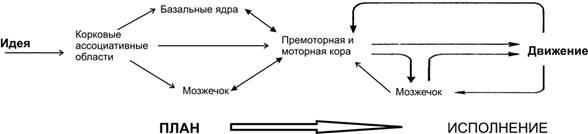
Рис. 14–1. Общая схема контроля произвольных движений.
Команды для произвольного движения исходят из ассоциативной области коры. В планировании принимают участие базальные ядра и латеральная часть мозжечка. Эти же структуры через таламус передают информацию премоторной и моторной областям коры. Моторная кора посылает сигналы по кортикоспинальному и кортикобульбарному трактам (корково–спинномозговые и корково-ядерные пути к двигательным нейронам мозгового ствола и спинного мозга). Движения вносят коррективы в сенсорные посылки от мышц, сухожилий и суставов, которые посредством обратной связи поступают к коре больших полушарий и мозжечку. Из мозжечка информация поступает в ствол мозга, где проецируется на пути, имеющие отношение к позе и координации: красноядерно–спинномозговой, бульборетикуло–спинномозговой, крыше–спинномозговой и пpеддвеpно–спинномозговой тракты. Поза постоянно корригируется не только перед началом, но и во время движения: движения сглаживаются и координируются мозжечком и его связями. Базальные ядра и латеральная часть мозжечка имеют обратную связь с премоторной и моторной корой, участвуя в планировании и организации произвольного движения.
Кортикоспинальная и кортикобульбарная системы Моторная кора
В моторной коре различают первичную моторную кору, а также премоторную и дополнительную моторную области (рис. 14–2А).
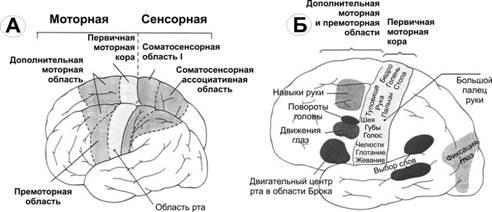
Рис.14–2.Двигательнаякорабольшогомозга.А. Моторная и соматосенсорная функциональные области. В первичной моторной коре представлены сверху вниз (на рисунке) области тела: от стопы до головы.Б. Представительство различных мышц в моторной коре и локализация корковых областей, отвечающих за специальные движения.
Первичнаямоторнаякора. На рис. 14–2А дана грубая топография представительства различных мышц начиная от мышц лица (вблизи от сильвиевой борозды) и кончая мышцами ноги (область в глубине продольной щели). В первичной моторной коре картированы участки, стимуляция которых вызывает сокращения отдельных мышц, но чаще возбуждаются мышечные группы.
Премоторнаяобластьрасполагается кпереди от первичной моторной коры, её топографическая организация подобна организации первичной коры: наиболее латерально располагаются зоны лица и рта, далее вверх к продольной щели — кисть, рука, туловище и нижние конечности. В премоторной области генерируются сложные спектры движений (например, движения плеча, руки, особенно кисти).
Кисть точно ориентирована для выполнения специфической работы. Чтобы достигнуть такого результата, передняя часть премоторной области формирует «двигательный образ» всех мышечных движений, которые должны осуществляться. Затем этот образ возбуждает каждый последовательный спектр мышечной активности в задней премоторной коре, требующийся для достижения «образа». Задняя премоторная кора посылает сигналы либо непосредственно в первичную моторную кору, возбуждая в итоге различные группы мышц, либо (чаще) через базальные ядра к первичной коре через таламус. Такимобразом,премоторнаякора,базальныеядра,таламусобразуюткомплекснуюсистемуконтроляикоординацииспектрамышечнойактивноститела.
Дополнительнаямоторнаяобластьрасполагается в продольной щели и функционирует совместно с премоторной областью, обеспечивая движения, поддерживающие осанку, фиксацию движений различных сегментов тела, позиционные движения головы и глаз и базу для тонкого моторного контроля кистей рук премоторной областью и первичной моторной корой.
Специализированныеобласти. На рис. 14–2Б показано несколько относящихся к двигательной функции специализированных центров (например, центры формирования речи, произвольного движения глаз, вращения головы, целевого движения кисти) коры больших полушарий, впервые обнаруженных нейрохирургами (Пенфилди Джаспер).
Центрформированияречи. Непосредственно перед первичной моторной корой выше сильвиевой борозды располагается участок коры, называемый областью Брока. Повреждение этой области не лишает человека способности произносить звуки, но он теряет способность к осмысленному произнесению слов. В лучшем случае человек способен произносить слова «да» или «нет». Рядом расположенные корковые области управляют дыхательной активацией голосовых связок и движениями рта и языка во время разговора. Таким образом, формирование речи нейронами премоторной области является комплексным процессом, вовлекающим в активность прилежащие области коры.
Центрпроизвольногодвиженияглаз. Над областью Брока располагается участок, контролирующий произвольные движения глаз. Повреждение этого участка лишает человека способности смещать глаза в направлении различных объектов.
Центрвращенияголовынаходится рядом с центром, контролирующим движения глаз, этот центр направляет голову в сторону объектов.
Центрцелевогодвижениякистилежит кпереди от первичной моторной коры. Повреждение этого центра делает движения кисти некоординированными и бессмысленными (моторная апраксия).
Сенсорныевходывмоторнуюкору. Функции моторной коры контролируются множеством сигналов из соматосенсорной коры, других сенсорных систем (в частности, зрения и слуха) и иных источников. Основные нервные пути, входящие в моторную кору:афферентные волокна из соматосенсорной области теменной коры, передних отделов фронтальной коры, слуховой и зрительной коры;афферентные волокна, проходящие через мозолистое тело из противоположного полушария мозга;соматосенсорные волокна, идущие непосредственно из вентро-базальных таламических ядер (тактильные сигналы и сигналы из мышц и суставов);нервные волокна вентролатерального и вентромедиального ядер таламуса, передающие информацию из мозжечка и базальных ядер;аксоны внутрипластинчатых ядер таламуса, контролирующие уровень возбудимости нейронов моторной коры.
Организациямоторнойкоры. Моторная кора организована в вертикальные колонки диаметром около 1 мм, в каждой из которых находятся тысячи нейронов, расположенных в 6 слоях коры. Отдельная колонка действует какфункциональнаяединица, стимулируя группу синергичных мышц и иногда отдельные мышцы.
Динамичныеистатичныенейронымоторнойкоры. Нейроны в любой отдельной колонке функционируют какинтегративныйпроцессор, используя информацию из различных источников. Вдобавок, каждая колонка может работать какусилитель, стимулируя большое количество путей, направляющихся к определённой мышце и одновременно к синергичным мышцам. Стимуляция одной пирамидной клетки не может возбудить мышцу. Чтобы вызвать мышечное сокращение обычно требуется одновременное возбуждение от 50 до 100 пирамидных нейронов. В каждой колонке имеются две группы пирамидных клеток: динамичные и статичные нейроны.Динамичныенейроныкратковременно и интенсивно возбуждаются в начале сокращения, вызывая начальное быстрое нарастание силы сокращения. Затем возбуждаютсястатичныенейроны, они продолжают удерживать силу сокращения в течение всего периода сокращения. Нейроны красного ядра (см. ниже) имеют подобные же динамические и статические характеристики, но отличаются тем, что в нём содержится большее количество динамичных нейронов, а в первичной моторной коре имеется больше статичных нейронов.
Сигналымоторнойкорывызывают специфические рефлекторные ответы спинного мозга. Например, когда из соответствующих отделов мозга поступает сигнал, возбуждающий мышцу, то моторной коре нет необходимости посылать сигналы для расслабления мышц–антагонистов. Это будет сделано системой реципрокной иннервации спинного мозга. Рефлексы растяжения функционируют постоянно, помогая демпфировать движения, вызванные сигналами из мозга, и усиливая их за счёт активации мышечных веретён. Другие рефлекторные механизмы спинного мозга (отдёргивание, шаг и ходьба, формирование и удержание позы) могут активироваться командами из головного мозга (см. табл. 13–2).
Обратнаясвязьмышечныхсокращенийсмоторнойкорой. Сокращения мышц, вызванные сигналами из моторной коры, посылают сигналы обратно от мышц в моторную кору. Основные соматосенсорные сигналы поступают из мышечных веретён, сухожильных органовГольджи, тактильных рецепторов кожи, покрывающей мышцы. Соматические сигналы усиливают мышечное сокращение посредством механизма обратной связи следующим образом. Если интрафузальные МВ сокращаются сильнее, чем сокращаются экстрафузальные МВ, то центральная часть веретена растягивается и возбуждает рецептор. Сигналы от веретён стимулируют пирамидные клетки моторной коры, сообщая о недостаточной силе сокращения мышц. Пирамидные клетки усиливают возбуждение мышц, способствуя выравниванию их сокращения с сокращением веретён. Роль тактильных рецепторов кожи можно проиллюстрировать примером, когда пальцы руки сжимают объект, пытаясь ухватиться за него. Сдавление кожи возбуждает тактильные рецепторы, что может усиливать сокращение мышц и способствовать более прочному обхвату предмета.
Нарушениефункциймоторнойкоры. Функции моторной контролирующей системы нарушаются при инсульте. Инсульт возникает при разрыве артериального сосуда внутри мозга или при тромбозе артерии, снабжающей кровью мозг. Поражается не только первичная моторная кора, но и соседние области и более глубокие структуры мозга, в том числе базальные ядра. В этих случаях наблюдаетсягемипарез— мышечный спазм поражённых мышц на противоположной стороне тела (из-за перекреста моторных путей). Этот спазм не является результатом потери функций первичной моторной коры или блокады кортикоспинальных волокон спинного мозга. Спазм возникает вследствие повреждения дополнительных нервных путей, берущих начало из внепирамидных участков коры. Эти пути в нормальных условиях тормозят активность вестибулярных и ретикулярных ядер ствола головного мозга. Когда эти ядра растормаживаются, они становятся спонтанно активными и вызывают интенсивное повышение спастического тонуса мышц.