

gistologia
.pdfтягиваются в плоские, связанные между собой клетки и образуют эндотелиальную выстилку первичных кровеносных сосудов. В результате соединения разрозненных вначале островков обра зуется сосудистая сеть желточного мешка.
Часть стволовых клеток превращается в крупные базофильные бластные клетки — п е р в и ч н ы е к р о в я н ы е к л е т к и . Ин тенсивно размножаясь внутри сосуда, большая часть первичных кровяных клеток постепенно утрачивает базофилию, все сильнее окрашиваясь кислыми красителями. Это происходит в связи с синтезом и накоплением в цитоплазме гемоглобина. Одновременно
в |
ядре |
увеличивается количество конденсированного |
хроматина. |
||
Такие |
клетки |
называют |
п е р в и ч н ы м и э р и т р о б л а с т а м и . |
||
В |
некоторых |
первичных |
эритробластах распадается и |
исчезает |
|
ядро. Образующаяся генерация ядерных и безъядерных первич ных эритроцитов разнообразна по размерам, однако чаще всего
встречаются крупные клетки |
— |
м е г а л о б л а с т ы и м е т а л о - |
ц и т ы . Мегалобластический |
тип |
кроветворения характерен для |
эмбрионального периода. При некоторых формах анемий (В^-де- фицитные, фолиеводефицитные и др.) в крови взрослых животных появляются клетки, сходные с первичными мегалоцитами.
Часть первичных кровяных клеток в эмбриональных сосудах проходит более длительный путь развития и образует популяцию вторичных эритроцитов, имеющих меньшие размеры и по внешне му виду мало отличающихся от эритроцитов взрослых животных.
Одновременно с внутрисосудистым эритропоэзом из первичных кровяных клеток вне сосудов развивается небольшое количество гранулоцитов — нейтрофилов и эозинофилов, т. е. происходит миелопоэз.
Возникшие в желточном мешке стволовые клетки с кровью переносятся в различные формирующиеся органы организма, где и происходит их дальнейшее развитие в разнообразные клетки крови. После закладки печени она становится универсальным ор ганом кроветворения; в ней экстраваскулярпо развиваются вторич ные эритроциты, зернистые лейкоциты и. мегакариоциты. К концу внутриутробного периода кроветворение в печени прекращается.
Из стволовых клеток в развивающемся тимусе (на 7—8-й не деле у крупного рогатого скота) дифференцируются лимфоциты тимуса и мигрирующие из него Т-лимфоциты. Последние заселя ют Т-зоны развивающихся периферических кроветворных орга нов — селезенки, лимфатических узлов. В начале своего развития селезенка также является органом, в котором образуются все виды форменных элементов крови. У большинства животных к концу эмбрионального периода в связи с массовым вселением Т- и В- лимфоцитов из центральных органов в селезенке и лимфатиче ских узлах начинает преобладать лимфоцитопоэз.
На поздних стадиях эмбрионального развития у животных ос новные кроветворные функции (роль центрального органа) начи нает выполнять красный костный мозг; в нем образуются эритро циты, гранулоциты, кровяные пластинки, часть лимфоцитов
(В-лимфоциты). В красном костном мозге сохраняется основная масса самоподдерживающей популяции стволовых клеток, и он становится органом, где происходит универсальный гемопоэз в постэмбриональный период. Таким образом, кроветворение, при сущее всей мезенхиме, становится функцией специализированных органов (печени, селезенки, тимуса, красного костного мозга, лим фатических узлов), где создаются соответствующие условия мик роокружения и кровотока.
Во время эмбрионального эритроцитопоэза идет характерпый процесс смены генераций эритроцитов, отличающихся морфологи ей и типом образующегося гемоглобина. Популяция первичных эритроцитов образует эмбриональный тип гемоглобина (Hb—F). На последующих стадиях эритроциты в печени и селезенке содер жат плодный (фетальный) тип гемоглобина (НЬ—Г). Наконец, на смену им появляется дефинитивный тип эритроцитов, которые образуются в красном костном мозге, они содержат третий тип гемоглобина (НЬ—А и НЬ—Аг). Разные типы гемоглобинов отли чаются составом аминокислот в белковой части.
СОЕДИНИТЕЛЬНЫЕ ТКАНИ
Широко распространенные в организме животных ткани с сильно развитой в межклеточном веществе системой волокон, благодаря которым эти ткани выполняют разносторонние механические и формообразующие функции — формируют комплекс перегородок, трабекул или прослоек внутри органов, входят в состав многочис ленных оболочек, образуют капсулы, связки, фасции и сухожи лия.
В зависимости от количественного соотношения между компо нентами межклеточного вещества — волокнами и основным веще ством и в соответствии с типом волокон различают три вида соеди нительных тканей: рыхлую соединительную ткань, для которой характерно количественное преобладание основного вещества пад комплексом разнообразно ориентированных и рыхло расположен ных коллагеновых и эластических волокон; плотную соединитель ную ткань, в ней резко выражено преобладание волокон над основ ным веществом, и ретикулярную ткань, содержащую в своем составе специфические ретикулярные волокна.
Основными клетками, создающими вещества, необходимые для построения волокон в рыхлой и плотной соединительной ткани, являются фибробласты, в ретикулярной ткани — ретикулярные клетки. Рыхлая соединительная ткань отличается особенно боль шим разнообразием клеточного состава.
РЫХЛАЯ СОЕДИНИТЕЛЬНАЯ ТКАНЬ
В организме рыхлая соединительная ткань является наиболее распространенной. Об этом свидетельствует тот факт, что она в большем или меньшем количестве сопровождает все кровеносные
144 |
Ю Заказ № 908 |
145 |
|
|

и лимфатические сосуды, формирует многочисленные прослойки внутри органов, входит в состав кожи и слизистых оболочек внут ренних полостных органов.
Независимо от локализации рыхлая соединительная ткань ^состоит из разнообразных клеток и межклеточного вещества, со держащего основное (аморфное) вещество и систему коллагеновых и эластических волокон. В соответствии с местными условия ми развития и функционирования количественное соотношение между этими тремя структурными элементами в различных участ-
.ках неодинаковое, что обусловливает органные особенности рых лой соединительной ткани.
Среди разнообразных высокоспециализированных клеток в •составе данной ткани различают более оседлые клетки (фибро пласты — фиброциты, липоциты), развитие которых в процессе жлеточного обновления происходит из предшественников, располо женных в пределах самой рыхлой соединительной ткани. Непо средственными предшественниками других более подвижных кле- ITOK (гистиоциты — макрофаги, тканевые базофилы, плазмоциты) являются клетки крови, активная фаза функционирования кото рых осуществляется в составе рыхлой соединительной ткани. В совокупности все клетки рыхлой соединительной ткани пред оставляют единый диффузно рассредоточенный аппарат, находя- •щийся в неразрывной функциональной связи с клетками сосуди стой крови и лимфоидной системы организма.
Повсеместная распространенность рыхлой соединительной тка ни, разнообразие и большое количество клеточных элементов,
способных к размножению и |
миграции, обеспечивает |
главные |
.(функции этой соединительной |
ткани: т р о ф и ч е с к у ю |
(обмен |
ные процессы и регуляция питания клеток), з а щ и т н у ю (учас тие клеток в иммунных реакциях — фагоцитоз, выработка иммуно
глобулинов и |
других веществ) и |
п л а с т и ч е с к о ю (участие в |
.-восстановительных процессах при тканевом повреждении). |
||
К л е т к и . |
Адвентициальные |
клетки — вытянутые клетки |
^звездчатой формы с овальным ядром, богатым гетерохроматином. -Цитоплазма базофильна, содержит немногочисленные органеллы.
.Располагаются вдоль наружной поверхности стенки капилляров •'и являются относительно малодифференцированными клеточными элементами, способными к митотическому делению и превраще-
.нию в фибробласты, миофибробласты и липоциты (рис. 102).
Фибробласты (libra — волокно, blastos — росток, зачаток) — постоянные и наиболее многочисленные клетки всех видов соеди нительной ткани. Это основные клетки, принимающие непосред ственное участие в формировании межклеточных структур. Они •синтезируют и выделяют высокомолекулярные вещества, необхо димые как для построения волокон, так и для образования аморф ного компонента ткани. В ходе зародышевого развития фиброблас ты возникают непосредственно из мезенхимных клеток. В пост- -эмбриональном периоде и при регенерации основными : предшественниками фибробластов являются адвентициальные
Рис. 102. Рыхлая соединительная ткань подкожной клетчатки кролика (до Ясвоину):
1 — эндотелий капилляра; 2 — адвентициальная камбиальная клетка; 3 — фибробласты; 4 — гистиоцит; 5 — жи ровая клетка.
клетки. Кроме того, молодые формы этих клеток сохраняют способность размножаться пу тем митотического деления.
По степени зрелости и, сле довательно, по структурной ха рактеристике и функциональной
активности различают три разновидности фибробластов. Мало- д и ф ф е р е н ц и р о в а н н ы е фибробласты имеют вытянутую, веретеновидную форму с немногочисленными короткими отросткамиОвальное ядро имеет хорошо выраженное ядрышко. Цитоплазма при световой микроскопии препаратов, окрашенных основными красителями, базофильная. При электронной микроскопии в цито плазме обнаруживают много свободных полисом и лишь короткие? узкие канальца гранулярной сети. Элементы комплекса Гольджи. расположены в околоядерной зоне. Митохондрии немногочисленны, и имеют плотный матрикс. Считают, что такие незрелые клетки обладают низким уровнем синтеза специфических белков. Их. функция сводится к синтезу и секреции гликозаминогликанов.
З р е л ы е ф и б р о б л а с т ы при рассматривании сверху — крупные (диаметр до 50 мкм) отростчатые клетки, содержащиесветлые овальные ядра с 1—2 крупными ядрышками и значитель ный объем слабобазофильной цитоплазмы. Периферическая зона клетки окрашивается особенно слабо, вследствие чего контуры еепочти незаметны. В поперечном разрезе уплощенное тело клеткп веретеновидной формы, так как его центральная часть, содержа щая ядро, значительно утолщена. Электронно-микроскопическж для цитоплазмы зрелого фибробласта характерна развитая грану лярная эидоплазматическая сеть, состоящая из удлиненных и рас ширенных цистерн, к мембранам которых прикреплены полисо мы больших размеров. Хорошо выражены и элементы (цистерны,. микро- и макропузырьки) комплекса Гольджи, распределенные повсей цитоплазме. Обнаруживаются различные по форме и вели чине митохондрии (рис. 103).
В функциональном отношении зрелые фибробласты являются клетками со сложной синтетической и секреторной деятельностью. Они одновременно синтезируют и выводят несколько типов спе цифических белков (проколлаген, проэластин, ферментные белки) и разнообразные гликозамипогликаиы. Наиболее отчетливо выра жена способность к синтезу белка коллагеновых волокон. На
•146 |
10* |
Н7 |

Рис. 103. Электронная микрограмма участка фибробласта (по Радостиной):
I — ядро; г — гранулярная эндоплазматическая сеть; в — митохондрии.
полисомах гранулярной сети
синтезируются а-цепи |
белка, а |
в полости компонентов |
ретику- |
лума осуществляется их связы вание в тройную спираль моле кулы проколлагена. Последние с помощью микропузырьков пе реносятся в цистерны комплекса Гольджи и затем в составе сек реторных гранул выделяются из клетки. На поверхности фибро бласта от молекул проколлагена отделяются концевые неспирализованные пептидные участки, они превращаются в молекулы
тропоколлагена, которые, полимеризуясь, формируют коллагеновые микрофибриллы и фибриллы (рис. 104). Гликозаминогликаны об разуются в комплексе Гольджи. Накапливаясь между клетками, ови создают условия для концентрации и полимеризации молекул тропоколлагена, а также входят в состав фибрилл в качестве це ментирующего компонента.
Фибробласты обладают подвижностью. В периферической зоне цитоплазмы располагаются актинсодержащие микрофиламенты, при сокращении которых обеспечиваются образование выступов и движение клетки. Двигательная активность фибробластов усили вается в регенераторную фазу воспалительной реакции при обра зовании соединительнотканной капсулы.
В грануляционной ткани появляются фибробласты с большим количеством сократимых нитей — м и о ф и б р о б л а с т ы , способ ствующие закрытию раны.
Волокнообразование в соединительной ткани приводит к тому, что часть фибробластов оказывается заключенной между близко расположенными волокнами. Такие клетки называют ф и б р о ц и т а м и . Они утрачивают способность к делению, принимают силь но вытянутую форму, у них уменьшается объем цитоплазмы и значительно снижается синтетическая активность.
Гистиоциты (макрофаги) в составе широко распространенной соединительной ткани являются наиболее многочисленной груп пой свободных, способных к миграции клеток, относящихся к си стеме моноиуклеарных фагоцитов (СМФ). В соединительноткан ных прослойках разных органов их количество неодинаковое и, как правило, значительно увеличивается при воспалении.
Рис. 104. Схема образования коллагеновой фибриллы:
При световой микроскопии окрашенных пленочных препаратов гистиоциты имеют разнообразную форму и разную величину (10—50 мкм), располагаются одиночно или группами. В сравне нии с фибробластами выделяются более очерченными, но неров ными границами и интенсивно окрашенной цитоплазмой, в кото рой имеются вакуоли, и включения. Ядро небольшое, овальное, слегка вогнутое, содержит много глыбок гетерохроматина, в свя зи с чем оно более темное (рис. 105).
Электронно-микроскопически на плазмолемме отмечают микро ворсинки, псевдоподии, инвагинации. В цитоплазме присутствует значительное количество лизосом, фагосом, гранул и липидных включений. Гранулярная сеть почти не развита. Митохондрии и комплекс Гольджи более развиты в активированных макрофагах. Цитохимическими методами в цитоплазме гистиоцитов выявляют разнообразные ферменты (кислые гидролазы, изоферменты кис лой фосфатазы, эстеразы и др.), с помощью которых происходит переваривание поглощенных веществ.
П о н я т и е о с и с т е м е м о н о и у к л е а р н ы х ф а г о ц и т о в (макрофагической системе). К этой системе относят находя-
149
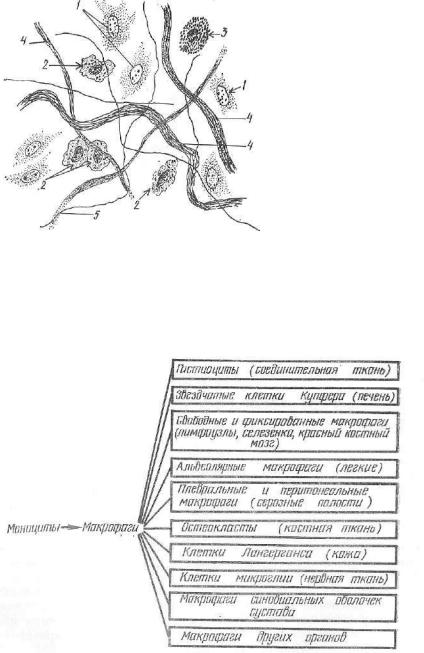
Рис. 105. Рыхлая соедини тельная ткань в пленочном препарате:
1 — фибробласты; 2 — гистио циты; з — тканевый бавофип; 4 — коллагеновые волокна; 5 — эластические волокна.
щиеся во многих тканях и органах клетки, обла дающие интенсивным эндоцитозом (фагоцито зом и пиноцитозом) мак ромолекул ярных ве ществ экзогенной и эн догенной природы, ча стиц, микроорганизмов,, вирусов, клеток, продук
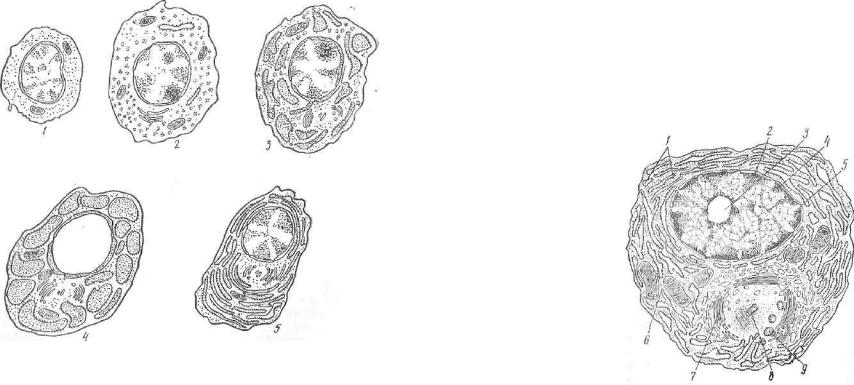
тов клеточного распада и т. п. Все макрофаги независимо от ло кализации происходят из стволовой кроветворной клетки краспога костного мозга, а их непосредственными предшественниками явля ются моноциты периферической крови. Моноциты, вышедшие из сосуда и попавшие в соответствующее микроокружение, адапти руются к новой среде и превращаются в органо- и тканевоспецифические макрофаги (рис. 106).
Рис. 106. Разновидности клеток, принадлежащих к системе мононуклеарных фагоцитов — СМФ (по Ван-Фюрту, 1980).
150
Таким образом, моноциты циркулирующей крови представляют -подвижную популяцию относительно незрелых клеток будущих зрелых макрофагов на их пути от костного мозга в органы и тка ни. В условиях культивирования макрофаги способны прочно прикрепляться к поверхности стекла и приобретать уплощенную форму.
В зависимости от локализации (печень, легкие, брюшная по-
.лость и т. п.) макрофаги приобретают некоторые специфические черты строения и свойства, позволяющие отличать их друг от
.друга, однако всем им свойственны некоторые общие структур ные, ультраструктурные и цитохимические признаки. Благодаря наличию сократимых микрофиламентов, обеспечивающих подвиж ность плазмолеммы, клетки этой системы способны к образованию различных приспособлений (ворсинок, псевдоподий, выступов), облегчающих захватывание частиц. Один из основных ультра структурных признаков макрофагов — наличие в их цитоплазме многочисленных лизосом и фагосом. С участием лизосомальных ферментов (фосфатазы, эстеразы и др.) фагоцитировапный мате риал расщепляется и перерабатывается.
Макрофаги — многофункциональные клетки. Основоположни ком учения о цитофизиологии клеток макрофагической системы является И. И. Мечников. До настоящего времени актуальны мно гие положения, сформулированные им о механизмах фагоцитоза и биологическом значении этого явления. Макрофагическая систе ма благодаря способности ее клеток поглощать и переваривать
.различные продукты экзо- и эндогенного происхождения пред ставляет одну из важнейших защитных систем, участвующих в •поддержании стабильности внутренней среды организма.
Макрофагам отведена важная роль в осуществлении защитной воспалительной реакции. Обладая способностью к направленному движению, определяемому хемотаксическими факторами (вещест ва, выделяемые бактериями и вирусами, иммунные комплексы •антиген—антитело, продукты тканевого распада, медиаторы лим фоцитов и др.), макрофаги мигрируют в очаг воспаления и стано вятся доминирующими клетками хронического воспаления. При этом они не только очищают очаг от инородных частиц и разру шенных клеток, но и стимулируют в последующем функциональ ную активность фибробластов. При наличии в очаге токсичных и устойчивых раздражителей (некоторые микроорганизмы, химиче ские вещества, малорастворимые материалы) с участием макро фагов формируется гранулема, в которой путем слияния клеток могут образовываться гигантские многоядерные клетки.
Макрофаги имеют существенное значение во многих иммуно логических реакциях: в распознавании антигена, его переработке •ж предъявлении лимфоцитам, в межклеточном взаимодействии с Т- и В-лимфоцитами и в выполнении эффекторных функций.
На поверхности плазмолеммы макрофага расположено два типа специфических рецепторов: рецепторы для Fc-части иммуно глобулинов и рецепторы для комплемента, особенно для его ком-
151
понента Сз. Поэтому в фазе распознавания и поглощения большое значение имеет опсонизация антигенов, то есть предварительное присоединение к ним иммуноглобулинов или комплекса иммуно глобулина с комплементом. Последующее прикрепление таких сенсибилизированных антигенов (иммунных комплексов) к соот ветствующим рецепторам макрофага вызывает движение псевдо подий и поглощение объекта фагоцитоза. Существуют также неспецифические рецепторы, благодаря которым клетка может фагоцитировать денатурированные белки или индифферентные частицы (полистирол, пыль и т. п.). С помощью пиноцитоза мак рофаги способны распознавать и поглощать растворимые антиге ны (глобулярные белки и др.).
Большая часть поглощенных антигенных веществ во многих фагоцитах разрушается полностью. Такая функция ликвидации избытка антигенов, проникших во внутреннюю среду организма, характерна для макрофагов печени, синусов селезенки, мозгового вещества лимфатических узлов. Особые разновидности специали зированных макрофагов — отростчатые «дендритные» клетки В-зон и «интердигитирующие» клетки Т-зон лимфатических узлов и селезенки. На поверхности их многочисленных отростков кон центрируются и сохраняются исходные или частично перерабо танные иммуногенные антигены. В этих зонах макрофаги вступа ют в кооперативное взаимодействие с В- и Т-лимфоцитами при развитии как гуморального, так и клеточного иммунитета.
Эффекторная функция макрофагов выражается не только в обезвреживании антигенов, но и в способности вырабатывать и выделять растворимые факторы, стимулирующие пролиферацию и дифференциацию Т- и В-лимфоцитов и регулирующие их актив ность. В некоторых случаях макрофаги могут быть эффекторными клетками в цитотоксических реакциях. Считают, что это важ ный механизм разрушения клеток с внутриклеточно паразитиру ющими микроорганизмами и опухолевых клеток.
Клетки системы монопуклеарных фагоцитов — активные участ ники миелоидного и лимфоидного кроветворения. Макрофаги в красном костном мозге являются своеобразными центрами, вокруг которых группируются предшественники развивающихся эритро цитов. Эти макрофаги участвуют в передаче накапливаемого же леза в клетки эритроидного ряда, поглощают ядра нормоцитов и фагоцитируют повреяеденные и старые эритроциты. Другие мак рофаги костного мозга фагоцитируют части мегакариоцитов после отделения от них кровяных пластинок. С помощью макрофагов селезенки происходит интенсивный эритрофагоцитоз и поглоще ние стареющих кровяных пластинок, а макрофагов всех лимфоидных органов — фагоцитоз плазмоцитов и лимфоцитов.
Тканевые базофилы (лаброциты, тучные клетки) обнаружены у большинства позвоночных и у всех млекопитающих, однако ко личество их у животных разных видов и в соединительной ткани различных органов неодинаковое. У некоторых животных отмеча ют обратно пропорциональную зависимость .между количеством
тканевых базофилов и базофилов крови, что свидетельствует о сходном биологическом значении этих типов клеток в системе тканей внутренней среды (например, у морских свинок тканевых базофилов много, но мало базофилов крови). Значительное коли чество тканевых базофилов содержится в подэпителиальной соеди нительной ткани кожи, пищеварительного тракта, дыхательных путей, матки. Их обнаруживают в соединительнотканных прослой ках по ходу мелких кровеносных сосудов в печени, почках, эндо кринных органах, молочной железе и других органах.
По форме тканевые базофилы чаще овальные или шаровидные, размером от 10 до 25 мкм. Ядро расположено центрально, в нем много глыбок конденсированного хроматина. Наиболее характер ная структурная особенность тканевых базофилов — наличие мно гочисленных крупных (0,3—1 мкм) специфических гранул, рав номерно заполняющих большую часть объема цитоплазмы и окрашивающихся метахроматически. Электронно-микроскопически в цитоплазме обнаруживают немногочисленные митохондрии, по лисомы и рибосомы. Эндоплазматическая сеть и комплекс Гольджи развиты слабо. На плазмолемме пальцевидные выпячивания. Специфические гранулы окружены мембраной и имеют неодина ковую электронную плотность; в некоторых гранулах содержатся еще более электроноплотные зерна или пластинки.
Характерное метахроматическое окрашивание гранул обуслов лено наличием в них сульфатированного гликозаминогликана — гепарина. Кроме того, в гранулах тканевых базофилов содержатся важнейшие биологические амины — гистамип, серотонин, дофа мин, имеющие многообразное фармакологическое действие. Цито химическими методами в цитоплазме выявлены различные фер менты — кислая и щелочная фосфатазы, липаза. Гистамин образу ется из аминокислоты гистидина с помощью гястидиндекарбоксллазы, которая является маркерным ферментом для тучных кле ток.
Располагаясь вблизи мелких кровеносных сосудов, тканевые базофилы одни из первых клеток реагируют на проникновение антигенов из крови. На их плазмолемме, так же как и у базофи лов крови, находится значительное количество иммуноглобулинов класса Е (IgE). Связывание антигенов и образование комплекса антиген—антитело сопровождается дегрануляцией и выделением из тканевых базофйлов^еосудистоактивиых веществ, обусловлива ющих появление местных и общих реакций. Гистамин повышает проницаемость стенки капилляров и основного вещества соедини тельной ткани, стимулирует миграцию эозинофилов, активирует макрофаги и т. д. Гепарин препятствует свертыванию крови. Уста новлено участие тканевых базофилов при развитии аллергических и анафилактических реакций.
Дегрануляция тканевых базофилов может быть вызвана и раз личными физическими факторами — травмой, резкими темпера турными воздействиями и др.
Плазмоциты (плазматические клетки) в функциональном отно-
152 |
153 |
|
шении — эффекторные клетки иммунологических реакций гумо рального типа, то есть реакций, сопровождающихся увеличение!* в крови циркулирующих антител, с помощью которых осуществ ляется обезвреживание антигенов, вызвавших их образование*- Это высокоспециализированные клетки организма, синтезирую щие и выделяющие основную массу разнообразных антител (им муноглобулинов).
По происхождению плазмоциты представляют конечные ста дии развития стимулированных антигеном В-лимфоцитов, которые* в местах своего расположения с участием клеток Т-хелперов ш макрофагов подвергаются активации, интенсивно размножаются. и превращаются в зрелые плазматические клетки. В наибольшем количестве плазмоциты находятся в селезенке, лимфатических узлах, в составе соединительной ткани слизистых оболочек пище варительного канала и дыхательных путей, в интерстпциалыюй соединительной ткани различных желез организма.
Развитие клеток плазматического ряда из стимулированных В-лимфоцитов происходит через стадии плазмобласта (иммунобласта), проплазмоцита и плазмоцита (рис. 107).
Рис. 107. Схема развития плазмоцитов (до Вайсу):
1 — предшественник плазмоцита (полустволовая клетка); 2 — плазмобласт; Я— юный плазмоцит; 4 — плазмоцит с расширенными цистернами эндоплазматической сети; 5 — зрелый плазмоцит.
П л а з м о б л а с т — крупная клетка (до 30 мкм в диаметре), ^имеющая светлое центрально расположенное ядро. В последнем обнаруживают небольшие гранулы расположенного по периферии хроматина и 1—2 ярко выраженных ядрышка. Электронно-мик- роскопически в цитоплазме выявляют очень редкие и мелкие цистерны гранулярной эндоплазматической сети, а также большое количество свободных полисом и рибосом. У немногочисленных митохондрий светлые матрикс и редкие кристы. Среди плазмо-
бластов |
часто встречаются митозы. П р о п л а з м о |
ц и т характе |
ризуется |
несколько меньшими размерами, резко |
выраженной |
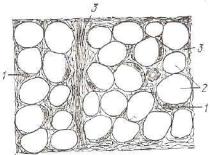
базофилией цитоплазмы и неровной из-за многочисленных выпя чиваний плазмолеммы поверхностью клетки. В цитоплазме нахо дится большое количество расширенных цистерн и мешочков гранулярной эндоплазматической сети. Между элементами грану лярной сети расположены небольшие по размеру митохондрии. Считают, что эти клетки могут вырабатывать и выделять иммуно глобулины. З р е л ы е п л а з м о ц и т ы — сравнительно неболь шие (8—10 мкм) клетки овальной формы с выраженными грани цами. В сильнобазофильной (пиронинофильной) цитоплазме об наруживают светлую околоядерную зону. Ядро округлое, распо ложено эксцентрично и содержит крупные глыбки гетерохроматиаа, распределенные в виде спиц колеса. Особенно характерно в «троении этих клеток при электронно-микроскопическом исследо вании наличие в цитоплазме многочисленных, расположенных -близко друг к другу длинных цистерн, имеющих очень узкую по лость и сближенные мембраны, на внешней поверхности которых находятся многочисленные полисомы. В околоядерной зоне с бо лее светлой цитоплазмой этих цистерн нет, в ней расположены центриоли и хорошо развитый комплекс Гольджи (рис. 108).
Таким образом, в ко нечной стадии развития плазмоциты содержат мощ ный белоксинтезирующий •аппарат, с помощью кото рого осуществляется синтез молекул иммуноглобулинов (антител). Установлено, что легкие цепи иммуно глобулинов синтезируются
Рис. 108. Схема ультрамикро скопического строения плазма тической клетки (по Бесси):
1 — гранулярная эндоплазматическая сеть; 2 — хроматин; 3 — ядрышко; 4 — ядерная оболочка; б — пора ядерной оболочки; в— •свободные рибосомы; 7 — комп лекс Гольджи; * — центриоли; & — секреторные пузырьки.
154 |
155 |
на полирибосомах гранулярной сети отдельно |
от тяжелых це |
пей. Последние отделяются от полирибосом |
после образования |
их комплекса с легкими цепями. Так как весь белоксинтезирующий механизм запрограммирован на синтез антител только одной разновидности, каждая плазматическая клетка определенного клона способна за один час синтезировать несколько тысяч моле кул иммуноглобулинов. Синтезированные молекулы поступают в просвет цистерн, а затем в комплекс Гольджи, откуда после при соединения углеводного компонента выводятся на поверхность клетки и выделяются. Выделение антител происходит и при раз рушении клетки.
Превращение В-лимфоцита в плазмоцит длится около суток; продолжительность активной антителопродуцирующей деятельно сти зрелых плазмоцитов составляет несколько дней. Зрелые плаз матические клетки не способны к делению, они стареют, погибают и поглощаются макрофагами.
Жировые клетки (липоциты) и жировая ткань (textus adiposus). Жировые клетки специализированы на синтезе и накаплива нии в цитоплазме запасных липидов, главным образом триглицеридов и утилизации их в соответствии с энергетическими и други ми потребностями организма. Липоциты широко распространены в рыхлой соединительной ткани и чаще располагаются не одиноч но, а небольшими группами по ходу мелких кровеносных сосудов. Во многих частях организма животных образуются значительные скопления жировых клеток, называемые жировой тканью. В эм бриогенезе жировые клетки возникают из клеток мезенхимы. Предшественниками для образования новых жировых клеток в постэмбриональный период являются адвентициальные клетки, сопровождающие кровеносные капилляры.
В связи с особенностями естественной окраски клеток, специ фикой их строения и функции, а также расположением различа ют у млекопитающих две разновидности жировых клеток и соот ветственно два типа жировой ткани: белую и бурую.
Б е л а я ж и р о в а я т к а н ь в организме животных разных видов и пород распределена неодинаково. В значительном коли честве она содержится в жировых депо: подкожная жировая ткань, особенно развитая у свиней, жировая ткань вокруг почек, в брыжейке, у некоторых пород овец у корня хвоста (курдюк). У животных мясных и мясо-молочных пород группы жировых клеток располагаются в перимизии и эндомизии внутри скелет ных мышц. Мясо, полученное от таких животных, обладает наи лучшими качествами («мраморное» мясо).
Структурная единица белой жировой ткани — шаровидные крупные (до 120 мкм в диаметре) зрелые жировые клетки с характерным микроскопическим строением (рис. 109). Большая часть объема клетки занята одной крупной каплей жира. Оваль но® ядро и цитоплазма находятся на периферии клетки. Такая клетка При овотошш микроскопии гистологического среза, окра- шоппого 0 примененном жирорастворяющих веществ, имеет перете
Рис. 109. Схема строения клеток беяой жировой ткани:
i — ядро жировой клетки; 2 — по лость, оставшаяся после растворе ния капли жира; з — соединитель ная ткань.
невидную форму. В результа те растворения жира на месте жировой капли в клетке оста ется светлая вакуоль. При электронной микроскопии в околоядерной зоне обнаружи
вают главным образом удлиненные митохондрии, другие органеллы слабо выражены. При развитии клеток жировые включения в цитоплазме появляются сначала в виде мелких рассеянных капель, позднее сливающихся в одну крупную каплю. Жировые вещества в клетках можно выявить при использовании специальных краси телей (судан III, судан IV, четырехокись осмия).
Из жировых клеток в жировой ткани образованы дольки раз личных размеров и формы. Между дольками находятся прослойки рыхлой соединительной ткани, в которых проходят мелкие крове носные сосуды и нервные волокна. Между жировыми клетками внутри долек располагаются отдельные клетки соединительной ткани (фиброциты, тканевые базофилы), сеть тонких аргирофильных волокон и кровеносные капилляры.
Общее количество белой жировой ткани в организме животных различных видов, пород, пола, возраста, упитанности колеблется от 1 до 30% к живой массе. Запасные жиры в жировой ткани — это наиболее высококалорийные вещества, при окислении которых в организме освобождается большое количество энергии (1 г жи ра =39 кДж). Подкожная жировая ткань, особенно у диких животных, имеет большое значение для защиты организма от механических повреждений, предохраняет от потерь тепла. Жи ровая ткань вдоль сосудисто-нервных пучков, в капсуле и оболоч ках органов обеспечивает их относительную изоляцию, защиту и ограничение подвижности. Скопления жировых клеток в сочета нии с окружающими их пучками коллагеповых волокои в коже подошв и лап создают хорошие амортизационные свойства. Зна чительна роль жировой ткани как депо воды. Образование воды— важная особенность обмена жиров у животных, обитающих в за сушливых районах (верблюды).
При голодании организм мобилизует прежде всего запасные жиры из клеток жировых депо. В них уменьшаются и исчезают жировые включения.
Жировая ткань главной орбиты, эпикарда, лап сохраняется да
же при сильном истощении.
Цвет жировой ткани зависит от вида, породы и типа кормле ния животных. У большинства животных, за исключением свп157-
ши
<яей и коз, в жире содержится пигмент каротин, придающий жел тый цвет жировой ткани.
Б у р а я ж и р о в а я т к а н ь в значительном количестве име ется у грызунов и животных, впадающих в зимнюю спячку, а также у новорожденных животных других видов. Расположена преимущественно под кожей между лопатками, в шейной области, в средостении и вдоль аорты.
Состоит она из относительно мелких клеток, очень плотно при бегающих друг к другу, напоминая внешне железистую ткань. К клеткам подходят многочисленные волокна симпатической нерв- <ной системы, они оплетены густой сетью кровеносных капилляspoB. Для клеток бурой жировой ткани в сравнении с клетками белой жировой ткани характерны центрально расположенное ядро и наличие в цитоплазме мелких жировых капель, слияния которых в более крупную каплю не происходит. В промежутках между жировыми каплями расположены многочисленные мито хондрии и значительное количество гранул гликогена. Содержа щиеся в митохондриях окрашенные белки системы транспорта электронов — цитохромы придают бурый цвет этой ткани.
В клетках бурой жировой ткани интенсивно идут окислитель ные процессы, сопровождаемые выделением значительного коли чества энергии. Однако большая часть образующейся энергии
.расходуется не на синтез молекул АТФ, а на теплообразование. Такое свойство липоцитов бурой ткани является важным для регуляции температуры у новорожденных животных и согревания животных после пробуждения от зимней спячки.
Пигментные клетки (пигментоциты), как правило, отростчатой формы. В цитоплазме много темно-коричневых или черных верен пигмента из группы меланинов. Значительное количество пигментных клеток — хроматофоров в соединительной ткани кожи низших позвоночных — рептилий, амфибий, рыб, у которых они •обусловливают ту или иную окраску внешнего покрова и выпол няют защитную функцию. У млекопитающих пигментные клетки •сосредоточены преимущественно в соединительной ткани стенки -глазного яблока — склере, сосудистой и радужной оболочках, а также в ресничном теле.
М е ж к л е т о ч н о е в е щ е с т в о рыхлой соединительной ткаши составляет значительную ее часть. Представлено оно коллагеяовыми и эластическими волокнами, расположенными относитель но рыхло и беспорядочно, и основным (аморфным) веществом. •В межклеточном веществе осуществляются разнообразные фер ментативные обменные процессы, перемещение различных веществ -ш клеточных элементов, самосборка и перестройка волокон в соот ветствии с направлением действия механических факторов. В меж клеточном веществе расположены чувствительные нервпые окон чания, непрерывно посылающие в центральную нервную систему -сигналы о его состоянии.
Коллагеновые волокна — основные волокна, обеспечивающие •механическую прочность ткани. В рыхлой соединительной ткани
они имеют вид лентовидных тяжей, ориентированных в различ ных направлениях. Волокна не ветвятся, им свойственна малая? . растяжимость, большая прочность на разрыв (выдерживают до* 6 кг на 1 мм2 поперечного сечения), способность объединяться в= пучки. При длительной варке коллагеновые волокна образуютклей (колла), откуда и произошло название волокон.
Прочность коллагеновых волокон обусловлена их более тон кой структурной организацией. Каждое волокно состоит из фиб рилл диаметром до 100 нм, расположенных параллельно другдругу и погруженных в межфибриллярное вещество, содержащеегликопротеиды, гликозаминогликаны и протеогликаны. Под элек тронным микроскопом по длине фибриллы наблюдают характер ную поперечную полосчатость — чередование темных и светлых полос с определенным периодом повторяемости, а именно один темный и один светлый сегмент вместе составляют один период?, длиной 64—70 нм. Наиболее отчетливо эта полосчатость видна на негативно окрашенных препаратах коллагеновых фибрилл. При электронной микроскопии позитивно окрашенных фибрилл, кроме основной темно-светлой периодичности, выявляют сложный рису нок более тонких электроноплотных полосок, разделенных узкими» промежутками шириной 3—4 нм.
В настоящее время характерный рисунок строения коллагеновой фибриллы объясняют специфичностью ее макромолекулярной? организации. Фибрилла состоит из более тонких микрофибрилл, образованных молекулами белка тропоколлагена. Последние име ют длину 280—300 нм и ширину 1,5 нм и являются своеобразны ми мономерами (рис. 110). Образование фибриллы — результат характерной группировки мономеров в продольном и поперечном? направлении. Мономеры уложены параллельными рядами и удер живаются друг около друга ковалентными поперечными связями,, причем в одном ряду между концами соседних мономеров имеет ся зазор, равный 0,4 длины периода, а по ширине мономеры одно го ряда накладываются на мономеры соседнего со смещением на lU его длины. Такое чередование зазоров и наложений создает полосчатый вид фибрилл на электронных микрофотограммах. Одна молекула тропоколлагена пересекает пять светлых п четыретемных сегментов (рис. 111).
Известно также, что по длине молекула тропоколлагена асим метрична и там, где сходные последовательности аминокислот оказываются друг против друга, возникают узкие вторичные темноокрашенные полосы. Каждая молекула тропоколлагена пред ставляет собой спираль из трех полипептидных цепей, удерживае мых друг около друга водородными связями. Уникальная структу
ра тропоколлагена обусловлена особенно высоким |
содержанием» |
в нем глицина (до 30%), а также оксилизина и |
оксипролина. |
В зависимости от аминокислотного состава и формы объединения: цепей в тройную спираль различают четыре основных тппа кол лагена, имеющих различную локализацию в организме. Коллаген? I типа является наиболее распространенным и содержится в со-
358 |
1593" |
Рис. 110. Схема строения коллаго- |
Рис. |
111. Коллагеновая фибрилла: |
|
|||||||
нового волокна: |
А — |
электронная микрофотография |
нега |
|||||||
|
|
|
||||||||
А — спиральная структура макро |
тивно |
окрашенной |
коллагеновой фибрил |
|||||||
молекулы коллагена (по Ричу); мел |
лы (ув. |
180 000); В — схема |
расположения |
|||||||
кие светлые |
кружки — глицин; |
тропоколлагеновых |
молекул, |
объясняющая |
||||||
крупные светлые кружки — пролин; |
возникновение |
поперечной |
исчерченности |
|||||||
заштрихованные кружки — гидрок- |
(по Ходжа и |
Петруски, |
1964): 1—темные |
|||||||
сипролин; В — схема строения кол- |
сегменты |
соответствуют |
промежуткам меж |
|||||||
лагеновых волокон; 1 — пучок фиб |
ду концами молекул |
тропоколлагена; |
2 — |
|||||||
рилл; 2 — |
фибрилла; 8 — протофи- |
светлые сегменты соответствуют зонам пе |
||||||||
брилла; |
4— |
молекула коллагена. |
рекрывания молекул. |
|
|
|
|
|||
единительной ткани кожи, сухожилий и в костях. Коллаген II ти па имеется преимущественно в гиалиновом и волокнистом хрящах. В коже зародышей, стенке кровеносных сосудов, связках преобла дает коллаген III типа, а в базальных мембранах — коллаген IV типа, в полипептидных цепях которого содержится особенно большое количество оксилизина.
Коллагеновые волокна неодинаковы по степени своей зрело сти. В составе новообразованных (при воспалительной реакции) волокон имеется значительное количество межфибриллярного це ментирующего полисахаридного вещества, которое способно вос станавливать серебро при обработке срезов солями серебра. Поэтому молодые коллагеновые волокна часто называют а р г и - р о ф и л ь н ы м и . В зрелых коллагеновых волокнах количество этого вещества уменьшается, и они утрачивают аргирофилию.
Эластические волокна имеют разную толщину (от 0,2 мкм в составе рыхлой соединительной ткани до 15 мкм в связках). На окрашенных гематоксилином и эозином пленочных препаратах соединительной ткани волокна представляют слабо выраженные тонкие ветвящиеся гомогенные нити, формирующие сеть. Для избирательного выявления эластических сетей используют специ альные красители — орсеин, резорцин — фуксин и др. В отличие от коллагеновых волокон эластические волокна не объединяются в пучки, обладают малой прочностью, высокой устойчивостью к воздействию кислот и щелочей, нагреванию и к гидролизующему действию ферментов (за исключением эластазы).
При электронной микроскопии в строении эластического во локна различают более прозрачную аморфную центральную часть, состоящую из белка эластина, и периферическую, в которой со держится большое количество электроноплотных микрофибрилл гликопротеидной природы, имеющих форму трубочек диаметром около 10 им. Последние вместе с межфибриллярным полисахаридным компонентом формируют футляр вокруг гомогенной части.
Образование эластических волокон в соединительной ткани обусловлено синтетической и секреторной функцией фибробластов. Считается, что вначале в непосредственной близости от фибробластов образуется каркас из микрофибрилл, а затем усилива ется образование аморфной части из предшественника эластина— проэластина. Под влиянием ферментов молекулы проэластина уко рачиваются и превращаются в небольшие, почти сферические молекулы тропоэластина. Последние при образовании эластина соединяются между собой с помощью уникальных веществ (десмозина и изодесмозина), отсутствующих в других белках. Кроме того, в эластине нет оксилизина и полярных боковых цепей, что обусловливает высокую устойчивость эластических волокон.
Особенно много эластических волокон в тех соединительно тканных образованиях, для которых характерны длительные напряжения и возвращение по окончании растяжения в перво начальное состояние (затылочно-шейная связка, брюшная желтая фасция). Высокая эластичность этих волокон в сочетании с отно сительной нерастяжимостью коллагеновых волокон создает гиб кую и прочную систему в соединительной ткани кожи и в стенках кровеносных сосудов.
Основное вещество. Все промежутки между клетками, волок нами и находящимися в рыхлой соединительной ткани сосудами микроциркуляторного русла заполнены бесструктурным основным веществом, которое на ранних стадиях развития ткани в количе ственном отношении преобладает над волокнами. В различных участках развитой соединительной ткани количество основного вещества неодинаково, значительное его содержание в подэпителиальных зонах соединительной ткани.
Основное вещество — гелеобразная масса, способная в широ ких пределах менять свою консистенцию, что существенно отра жается на его функциональных свойствах. По химическому соста ву это очень лабильный комплекс, состоящий из гликозаминогликанов, протеогликанов, гликопротеидов, воды и неорганических солей. Важнейшим химическим высокополимерным веществом в этом комплексе является несульфатированная разновидность гликозаминогликанов — гиалуроновая кислота. Неразветв ленные длинные цепи молекул гиалуроновой кислоты образуют много численные изгибы и формируют своеобразную молекулярную сеть, в ячеях и каналах которой находится и циркулирует ткане вая жидкость. Благодаря наличию таких молекулярных про странств в основном веществе имеются условия для передвиже ния различных веществ от кровеносных капилляров к клеткам
160 |
Ц Заказ № 90S |
161 |
|
|
соединительной и других тканей и продуктов клеточного метабо лизма в обратном направлении — к кровеносным и лимфатическим капиллярам для последующего их выделения из организма.
Образование основного вещества связано главным образом с двумя источниками: синтезом и выделением веществ из клеток (преимущественно из фибробластов) и поступлением их из крови. Поступающие в межклеточные пространства вещества подверга ются полимеризации. Полимеризованное или деполимеризованное состояние основного вещества является фактором, влияющим не только на связывание воды и транспорт содержащихся в тканевой жидкости растворимых компонентов (ионов, глюкозы, аминокис лот и др.), но и на миграцию клеток. Регулирующее влияние на состояние основного вещества оказывают многие гормоны (кортикостероиды и др.), действие которых направлено на клетки, а через них на компоненты межклеточного вещества. Под влиянием биогенных аминов и фермента гиалуронидазы происходит повы шение проницаемости основного вещества. Некоторые микроорга низмы, синтезируя и выделяя гиалуронидазу, вызывают деполи меризацию гиалуроновой кислоты основного вещества и таким способом ускоряют свое распространение в организме животного.
Для окрашивания основного вещества (гиалуроновой кислоты) используют основные красители, обладающие особенно высоким сродством к кислотным (анионным) участкам — например, альциановый синий или катионные метахроматические красители (то лу идиновый синий).
ПЛОТНАЯ СОЕДИНИТЕЛЬНАЯ ТКАНЬ
Этот вид соединительных тканей характеризуется количествен ным преобладанием волокон над основным веществом и клетка ми. В зависимости от взаимного расположения волокон и образо ванных из них пучков и сетей различают две основные разновид ности плотной соединительной ткани: неоформленную и оформ ленную.
В плотной неоформленной соединительной ткани волокна об разуют сложную систему перекрещивающихся пучков и сетей. Такое расположение их отражает разносторонность механических воздействий на данный участок ткани, соответственно которым и располагаются эти волокна, обеспечивая прочность всей тканевой системы. Плотная неоформленная ткань находится в большом количестве в составе кожного покрова животных, где она осущест вляет опорную функцию. Наряду с взаимопереплетающимися коллагеновыми волокнами в ней имеется сеть эластических волокон, обусловливающая способность тканевой системы к растяжению и возвращению в исходное состояние после прекращения действия внешнего механического фактора. Разновидности плотной не оформленной ткани входят в состав надхрящницы и надкостницы, оболочек и капсул многих органов.
162
упорядоченно располо женными волокнами, что соответствует действию меха
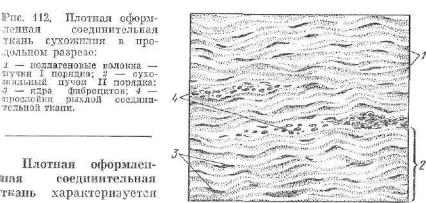
нического натяжения ткани в одном направлении. В соот ветствии с типом преобладающих волокон различают коллагеновую и эластическую плотные оформленные ткани. Плот н а я о ф о р м л е н н а я к о л л а г е н о в а я т к а н ь в наиболее типичном виде представлена в сухожилиях. Она состоит из плот но лежащих, параллельно ориентированных вдоль сухожилия коллагеновых волокон и сформированных из них пучков (рис. 112). Каждое коллагеновое волокно, состоящее из многочисленных фиб рилл, обозначают как пучок I порядка. Между волокнами (пуч ками I порядка), зажатые ими, расположены также продольно ориентированные фиброциты. Совокупность пучков I порядка об разуют пучки II порядка, окруженные тонкой прослойкой рыхлой соединительной ткани—эндотенонием. Несколько пучков II по рядка формируют пучок III порядка, окруженный более толстым слоем рыхлой соединительной ткани — перитенонием. В крупных сухожилиях могут быть и пучки IV порядка. Перитеноний и эндотеноний содержат кровеносные сосуды, питающие сухожилие, нервные окончания и волокна, посылающие в центральную первную систему сигналы о состоянии натяжения ткани.
П л о т н а я о ф о р м л е н н а я э л а с т и ч е с к а я т к а н ь у животных встречается в связках (например, в выйной). Она образована сетью толстых продольно вытянутых эластических волокон. В узких щелевидных пространствах между эластически ми волокнами расположены фиброциты и тонкие, переплетающие ся между собой коллагеиовые фибриллы. В некоторых местах имеются более широкие прослойки рыхлой соединительной ткани, по которым проходят кровеносные сосуды. Данная ткань, пред ставленная системой циркулярио расположенных мембран и эла стических сетей, имеется в крупных артериальных сосудах.
РЕТИКУЛЯРНАЯ ТКАНЬ
Эта ткань является разновидностью соединительных тканей, со стоит из отростчатых ретикулярных клеток и ретикулярных воло кон, формирующих трехмерную сеть (reticulum), в ячеях которой
И* |
163 |