

gistologia
.pdfИнтерфоза
Рис. 29. Митотический цикл. Схема (по Хему).
Центромера.
Ядрышко
Центраоль
Хромосомные
нити
Центральное деретено
Рис. 30. Схема митоза (по Манзия):
iciia и периферических, связанных с центромерами отдельных хро мосом. Хромосомы локализованы в этот период в экваториальной плоскости клетки и в совокупности образуют фигуру «материнской
.ик»;»ды», или «экваториальной пластинки» (д). В течение метафа йл сестринские хроматиды каждой хромосомы постепенно обособ- л Jпотея, сохраняя связь лишь в области центромера.
15 период а н а ф а з ы клетки вытягиваются по длинной оси
п\ I юматинового веретена и хроматиды хромосом, теряя связь друг
сдругом в области центромер, расходятся и становятся самостоя
тельными хромосомами формирующихся дочерних клеток (е, ж).
Т е л о ф а з а — конечная стадия митотического деления. В период телофазы токи цитоплазмы, характерные для этого периода митоза, обусловливают обособление двух дочерних клеток. Микро трубочки митотического аппарата материнской клетки разруша ются и исчезают. Хромосомы дочерних клеток включаются в про цессы синтеза рибонуклеиновых кислот, рассредоточиваются и микроскопически выявляются лишь частично как глыбки хромати на. Вновь образуется ядерная оболочка. По мере активизации «ндрышкового организатора» соответствующих хромосом форми руются ядрышки (з). По завершении процесса деления дочерние клотки переходят в период интерфазы.
Метафаза — наиболее благоприятный период клеточного цикла* дли изучения структурной организации хромосом. Установлено, что каждая хромосома представлена гигантской молекулой дезомипуклеопротеида (НДП), рассредоточенной в веществе ядра в период интерфазы и сложно уложенной и максимально уплотнен-: noii и период митотического деления. Для каждого вида животных кпрнкторно строго определенное количество хромосом. В частности. клотки крупного рогатого скота содержат 60 хромосом, лошади — (Ж, снииьи — 40, овцы — 54, собаки — 78.
Мри разнообразии размеров и форм митотические хромосомы продета ил яют собой палочковидные образования плотно упакован ных но отношению друг к другу двух хроматид. Каждая хромосо ма ра.чдоляотся первичной перетяжкой (центромерой, кинетохором) па дна плеча. Центромера в период митоза связана с микротруПочками неротеиа. В зависимости от расположения первичной Пвр^тмжки различают мота центрические хромосомы с равными Шмншмп, субмотацентрические хромосомы с различной длиной ШЮЧ и акроцонтрическио хромосомы, содержащие центромер в ипнци крпмоегшы (рис. 31). От-
ЛОЛМШв хромосомi.i имеют кто-
I'IIс иI. СХОМА общей морфологии
ХРОМОСОМ!
ммтпцоитричооких ('), оуЛмотпцшггрических (а), пкроцвитричооиих (твлоионтричогжих, я), спутничных (ядрышкопых, •!)', Т — теломеры; Ц — цонгромерш (порпичпые перетяжки), ПО — ядрышконмп организатор (вторичная пере тяжка).
48

ветствии с конечными этапами дифференцировки тканей, органа или системы органов в норме или в условиях патологии.
Морфологически амитоз характеризуется изменением формы и числа ядрышек с последующей перешнуровкой ядра. Образующие ся при этом двуядерные и многоядерные клетки при последующей цитотомии могут образовывать одноядерные клетки (рис. 33). По физиологическому значению различают три вида амитотического деления: амитоз генеративный, дегенеративный и реактивный. Г е- н е р а т и в н ы й а м и т о з — полноценное деление клеток, дочер ние клетки которых способны в последующем к митотическому делению и к характерному для них нормальному функционирова нию. Р е а к т и в н ы й м и т о з вызывается какими-либо неадекват
ными воздействиями на |
организм. Д е г е н е р а т и в н ы й |
ами |
т о з — деление, связанное |
с процессами дегенерации и |
гибели |
клеток. |
|
|
Г Л А В А 2 |
|
|
ОБЩАЯ ЭМБРИОЛОГИЯ |
|
|
Эмбриология (embryon — зародыш, logos — учение) — наука о раз витии зародыша. Она изучает индивидуальное развитие животных с момента зарождения (оплодотворения яйцеклетки) до его вылупления или рождения. В течение этого периода, для которого ха рактерна высокая степень формообразования и нарастания массы, одноклеточный организм приобретает-морфологическое сходство со взрослым животным. Эмбриология рассматривает развитие и строе ние половых клеток (гаметогенез) и основные этапы эмбриогене за: оплодотворение, дробление, гаструляцию, закладку осевых органов и органогенез, развитие провизорных (временных) орга нов. Эмбриология — это бурно развивающаяся отрасль биологиче ской науки, широко используемая современные методы исследова ния — электронную микроскопию, гистохимию, гистоавторадиографию, а также культуру тканей, микрохирургию и другие, что обусловило дальнейшее развитие эволюционной эмбриологии и ее главного направления — экспериментальной эмбриологии, созда ние которых связано с именами русских ученых К. Ф. Вольфа, К. М. Бэра, А. О. Ковалевского, И. И. Мечникова, А. Г. Кнорре, Г. А. Шмидта.
История эмбриологии, уходящая в годы далекой древности, — это история ожесточенной борьбы материалистического и идеали стического представления о мироздании.
Каждый этап эмбриогенеза является наглядным воплощением законов диалектического материализма: закона перехода количе ственных изменений в качественные, закона единства и борьбы противоположности, закона отрицания отрицания.
Эмбриология сельскохозяйственных животных изучает внутри утробное развитие зародышей в материнском организме или яйце. Эмбриогенез сельскохозяйственных животных является сложно
(протекающим процессом, в котором нашли отражение эволюция, «позвоночных (филогенез) и индивидуальное развитие (онтогенез).
Понять сложные вопросы эмбриогенеза сельскохозяйственных животных можно лишь после знакомства с основными чертами развития представителей разных классов позвоночных.
Достижения современной эмбриологии широко используют в животноводстве, птицеводстве, при разведении рыб; в ветеринарии л медицине при решении многих практических задач, касающих ся искусственного осеменения и оплодотворения, технологии уско ренного воспроизводства и селекции; повышения плодовитости 'Сельскохозяйственных животных, размножения животных путем трансплантации эмбрионов, генно-инженерных манипуляций с яйцеклетками и эмбрионами животных, при изучении патологии •беременности, при распознавании причин бесплодия и других itoupocoB акушерства. Способствуя формированию врачебного мыш ления, данные эмбриологии необходимы в повседневной практиче ской деятельности ветеринарного врача.
ПОЛОВЫЕ КЛЕТКИ (ГАМЕТОЦИТЫ)
31 о строению половые клетки сходны с соматическими (телесны ми) клетками. Они также состоят из ядра и цитоплазмы, постро енной из органелл и включений.
Отличительные свойства зрелых гаметоцитов — низкий уро вень процессов ассимиляции и диссимиляции, неспособность к де лению, содержание в ядрах гаплоидного (половины) числа хро мосом.
С т р о е н и е п о л о в ы х к л е т о к с а м ц о в . Половые клетки самцов (спермин) у всех позвоночных имеют жгутиковую форму (рис. 34).Образуются они в половых железах самцов (семенни ки v) в большом количестве. Так, в одной порции выделенного сеIVH4UJ (эякулята) у лошади,
собшш и других животным содержатся десятки миллио ном и даже миллиарды спермиои.
('термин сольскохозиистПОмгшх животных обладают подиижиостыо, на их форме отрижнптсн диш'птелышя актишни"п. клотки. Как размер, ТВИ И фирма пн'рмпон у раз-
PlIOi .'l'i. СпврМИИ домашних ЖИ
ВОТНЫХ!
1 — жороОца; 8 — осла; в — быка; 4 — Па liana; б — КОВЛА; б — хряка; Щ т кобеля; 8 — кота. Под 8, в, в—
ммд головки о профиль.
44 |
45 |
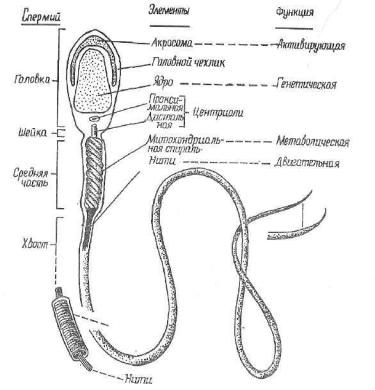
Рис. 35. Схема строения спермиев млекопитающих, на которой изображены структуры, выявляемые с помощью электронного микроскопа, и указаны выполняемые ими функции.
пых животных сильно варьируют. Строение этих клеток обуслов лено их функцией (оплодотворение половой клетки самки), по этому они состоят из головки, шейки и хвостового отдела ( рис. 35). Большую часть головки занимает плотное, богатое нуклеопр'отеидами ядро, в нем локализуется наследственный материал (мужской набор плотно упакованных хромосом). Спермин гетерогенны, так как в их ядрах содержатся разные типы половых хромосом. Поло вина спермиев имеет Х-хромосому, другая половина — У-хромо- сому. Половые хромосомы несут генетическую информацию, опре деляющую половые признаки самца. От остальных хромосом (аутосом) они отличаются большим содержанием гетер охроматина, размером и строением.
Передний край головки спермия покрыт цитоплазматическим чехликом, или а к р о с о м о й , которая образуется в результате сложной перестройки комплекса Гольджи. В акросоме находятся
46
ферменты. Под их влиянием при оплодотворении яйцеклетки разрушаются\ гликозаминогликаны (мукополисахариды), склеиваю щие клетки вторичной оболочки. Нарушение целостности вторич ной оболочки необходимо при проникновении спермия в яйцеклет ку. Акросома играет существенную роль в стимуляции яйцеклетки к дальнейшему развитию. Снаружи головка спермия покрыта плазмолеммой. Электронно-микроскопическими методами в ней найдены фибриллы, придающие ей механическую прочность. Не прерываясь, плазмолемма с головки спермия переходит на шейку, а затем хвостовой отдел.
За головкой расположена шейка, состоящая из двух центриолей: проксимальной и дистальной. Первая при оплодотворении вносится в цитоплазму яйцеклетки, обусловливая ее деление. Вто рая также имеет две части: передпюю и заднюю. От передней на чинается осевая нить хвостика, поэтому ее относят к аппарату движения спермия. Задняя часть дистальной центриоли имеет форму кольца и находится на границе начальной и главной частей хвостового отдела спермия.
Хвостовой отдел состоит из начальной, главной и концевой ча стей. В центре всего хвостового отдела проходит осевая нить. По строенная из микротрубочек, содержащих белок тубулин, она по строению сходна с мерцательными ресничками эпителиальных клеток. В начальной части хвостика сосредоточена основная масса цитоплазмы спермия. Здесь много митохондрий, которые выстраииаются по спирали вокруг осевой нити (рис. 36). Цитоплазма бога та гликогеном, фосфолипидами; ферменты сукцинатдегидрогеназа (СДГ), аденозинтрифосфатаза и другие характеризуются высокой активностью. Осевая нить главной части хвостика одета узким слоем цитоплазмы, лишенной органелл и включений. Снаружи расположена плазмолемма.
Концевая часть хвостового отдела состоит только из осевой нити и плазмолеммы.
Таким образом, в жгутиковых спермиях позвоночных последонятолыю расположены структуры, несущие генетическую, метабо лическую, двигательную функции (см. рис. 35). При оплодотворе нии спермий вносит в яйцеклетку отцовский генетический мате рин л и центросому (последняя отсутствует в яйцеклетке), нноПходимуго при делении зиготы (оплодотворенной яйцеклетки). Спермий обеспечивает встречу и создает необходимые условия для «иодроиин ого в яйцеклетку. Спермин способны к движению в на- «фшинчц'Ш яйцеклетки (хемотаксис) и против тока жидкости (роотдкоис). Они обладают минимальным запасом питательных пощэотп, которые очень быстро расходуются при движении клетки. Мели по проииондет слияния спермия с яйцеклеткой, то в половых путях самки он обычно погибает через 24—36 ч.
В искусственных условиях можно продлить жизнь мужских половых клеток до нескольких лет, применив глубокое охлажде ние, полностью снижающее обмен веществ клетки. Это свойство спермиев широко используется в настоящее время при искусст-
47

Рис. 36. Электронно-микроскопическое строение спермия:
1 — головка; 2 — шейка; 3 — осевая нить; 4 — митохондрии в связующем отделе; 5 — плазмолемма.
Яйцеклетки позвоночных овальной фирмы, неподвиж ны, состоят \из ядра и цито- к плазмы. Шаровидной формы п большого размера ядро по строено, как и у соматичес кой клетки, из хроматина, яд рышка, кариолимфы и кариолеммы. Ядро содержит гапло идный (одинарный) набор хромосом. Яйцеклетки млеко питающих относят к гомогаметным, так как в их ядре имеется только Х-хромосома. Ядрышко крупное. Его раз мер и строение свидетельству ет об интенсивном синтезе рибосом и РНК.
Для цитоплазмы характер на относительно высокая степень базофилии, обладающей срод
ством с РНК. В ней содержатся свободные рибосомы, эндоплазматическая сеть, комплекс Гольджи, митохондрии, мультивезикуляриыс тельца, желток. В образовании последнего активное участиепринимают эндоплазматическая сеть и комплекс Гольджи. Жел ток, являясь цитоплазматическим включением, имеет вид гранулг шаров и пластин; состоит из протеинов, углеводов, фосфолипидов.
Овоциты обладают полярностью. Это объясняется неравномер ным расположением желтка и других цитоплазматических струк тур, в связи с чем в овоците различают два полюса: апикальный? и Назальный.
Периферический слой цитоплазмы яйцеклетки называют к о р т и к а л ь н ы м с л о е м (cortex — кора). Он лишен полностью желтка, содержит множество митохондрий. Функции у этого' слоя — перепое питательных веществ и участие в развитии заро дыша на самых ранних стадиях эмбриогенеза.
Mii цок летки покрыты оболочками. Различают первичную, вто ричную и третичную оболочки. Первичная оболочка — это плазмоломми, Поэтому она присутствует всегда. Вторичная оболочка явлнотол производной фолликулярных клеток яичника. Она состоит ни фолликулярных клеток и выполняет трофическую, защитнуюФункции, препятствует полиспермии (оплодотворение яйцеклетки многими опормиями) и особенно хорошо развита у млекопитаю щих. Третичной оболочка овоцита формируется из материала, секритируомого клетками яйцевода. Она играет защитную, а такжетрофическую функции и развита у пресмыкающихся и птиц. Тре- 'шмиыми оболочками яйцеклетки птиц являются белок, подскорлуповая и скорлуповая оболочки яйца.
Таким образом, яйцеклетки в связи с выполняемыми ими функ циями обладают рядом морфологических особенностей: наличием'
А Заказ №. 908 |
49> |
запаса питательного материала (желтка), кортикального слоя, спе циальных оболочек, полярностью, генетической однородностью (Х-хромосомой).
Классификация яйцеклеток. Усложнение эмбриогенеза в ряду хордовых незначительно сказалось па строении половых клеток самцов, тогда как в яйцеклетках наблюдается постепенное нарас тание содержания питательных веществ (желтка). По количеству
желтка различают яйцеклетки |
с |
малым |
количеством желтка — |
о л и г о л е ц и т а л ь н ы е (oligos |
— |
мало, |
lecytos — желток), со |
средним количеством — м е з о л е ц и т а л ь н ы е (mesos — средний) и с большим количеством желтка — п о л и л е ц и т а л ь н ы е (po ly — много).
По месту расположения желтка в цитоплазме различают яйце
клетки с |
равномерным распределением желтка — и з о л е ц и - |
т а л ь н ы е , |
или г о м о л е ц и т а л ь н ы е (isos, homos — одинако |
вый, сходный), и с локализацией желтка у одного полюса — тело- л е ц и т а л ь н ы е (telos — край, конец). Олиголецитальные и изолецитальные яйцеклетки у ланцетника и млекопитающих. Если у ланцетника это связано с наличием в их развитии личиночной ста дии (личинка сама для себя добывает питательный материал), то у млекопитающих — в связи с внутриутробным развитием. Мезо лецитальные и телолецитальные яйцеклетки — у амфибий, неко торых рыб, а полилецитальные и телолецитальные — у многих рыб, пресмыкающихся, птиц и яйцекладущих млекопитающих.
РАЗВИТИЕ ПОЛОВЫХ КЛЕТОК (ГАМЕТОГЕНЕЗ)
Родоначальниками половых клеток являются первичные половые клетки — гаметобласты (гомобласты). Выявляются они микромор фологическими и цитохимическими методами в начале эмбриогене за в стенке желточного мешка (плодовая оболочка зародыша) вблизи кровеносных сосудов. Гонобласты — крупные клетки не правильно округлой формы с большим светлым ядром, цитоплазма богата гликогеном и характеризуется высокой активностью щелоч ной фосфатазы. Гонобласты интенсивно делятся митозом и с током крови или по ходу кровеносных сосудов мигрируют в зачатки по ловых желез (гонады), где окружаются поддерживающими (фол ликулярными) клетками. Последние выполняют трофическую функцию. Затем, в связи с развитием пола животного, половые клетки приобретают свойства, характерные для спермиев и яйце клеток.
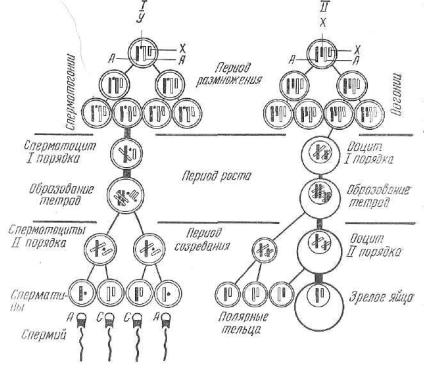
Р а з в и т и е с п е р м и е в (сперматогенез) (рис. 38—/) про текает в семенниках (половой железе самца) половозрелого жи вотного. В сперматогенезе различают четыре периода: размноже ние, рост, созревание и формирование.
Период размножения. На этой стадии клетки называются с п е р м а т о г о н и я м и . Они имеют небольшие размеры, диплоид ное (двойное) число хромосом, мелкие округлые ядра, богатые хроматином. Клетки интенсивно делятся митозом. Некоторые из
Л — аутосомы; X, Y — половые хромосомы. В период размножения показаны три поколения еперматогониев и оогониев, делящиеся митотически и остающиеся ди плоидными.
них перестают делиться, начинают увеличиваться в размере и пе реходят во второй период своего развития — период роста. Другая масть спермиев продолжает делиться. Они являются стволовымиклетками и пополняют запас сперматогоний.
Период роста. В этот период клетки называются п е р в и ч н ы ми с п е р м а т о ц и т а м и . У них сохраняется диплоидное число хромосом. В процессе роста увличивается размер клетки и проис ходят сложные изменения в перераспределении наследственного материала в ядре, в связи с чем различают четыре стадии: лептотонпую, зиготешгую, пахитенную, диплотенную.
Для л е п т о т е и п о й (leptos — тонкий, taenia — лента) стадии характерны хорошо выраженное ядрышко, ядерная тека и спира ли нация хромосом, котоые имеют вид тонких нитей. Гомологич ные хромосомы (отцовские и материнские) друг с другом не конъюгируют. При з и г о т е н н о й (zygoo — соединять), или синаптической (synapsis —соединение), стадии гомологичные хромосомы сближаются. Присоединившись друг к другу (конъюгировав)
50 |
4* |
5S |
|
|
« образовав пары — диады, обмениваются генами — кроссииговер (crossing — скрещивание, over — через). При п а х и т е н н о й (pachys — толстый) стадии пары гомологичных хромосом перекру чиваются друг с другом, сильно спирализуются, поэтому укорачи ваются и утолщаются. Д и п л о т е н н а я (diploos — двойной) ста дия — процесс формирования тетрад. В каждой паре возникает
.продольная щель, и в двух конъюгировавших хромосомах образу ются четыре хроматиды, которые начинают постепенно разъеди няться, сохраняя на некоторое время связь друг с другом в центре ювоими концами. Затем эти связи теряются.
Период созревания. Это процесс развития сперматид с поло винным числом хромосом, что достигается путем двух, следующих друг за другом делений созревания. После первого деления первич ного сперматоцита образуются два вторичных сперматоцита, каж дый из которых получает только половину от количества хромосом первичного сперматоцита. Вскоре за первым делением созревания без интерфазы (следовательно, без синтеза ДНК, характерного для периода S, в ходе которого увеличивается число хромосом) следует второе деление созревания. Из каждого вторичного сперматоцита образуется по две сперматиды. При этом пары хроматид, соединен ные между собой в центре центромерой, разъединяются и распре деляются по разным сперматидам, становясь хромосомами.
Следовательно, в процессе созревания из каждого первичного «сперматоцита возникает четыре сперматиды с одинарным числом хромосом. Половые Х- и У-хромосомы, характерные для спермиев, •расходятся по разным сперматидам, в связи с чем Х-хромосомами «снабжаются две сперматиды, другие две — У-хромосомами. Этот процесс обусловливает гетерогаметность самцов млекопитающих.
Способ деление, при котором дочерние клетки получают вдвое •меньше число хромосом, носит название м е й о з (meiosis — умень шение), или редукционное (reducere — уменьшение) деление.
Сперматиды имеют округлую форму, небольшие размеры, отно сительно крупное ядро, бедное хроматином. Митохондрии, комп лекс Гольджи, центросома хорошо развиты и расположены вблизи ядра. Другие органеллы, а также включения почти отсутствуют.
j Сперматиды не способны делиться.
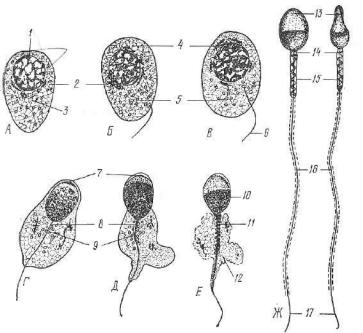
U" Период формирования (рис. 39). Завершающий период сперт матогенеза, в ходе которого округлая клетка — сперматида — при обретает морфологические свойства, характерные для спермия. В ходе формирования светлое округлое ядро сперматиды посте пенно уплотняется, становится овальным и перемещается к плазтдолемме. Комплекс Гольджи передвигается к ядру и продуцирует уплотеннную гранулу — а к р о б л а с т . Последний увеличивается в размере и в виде чехлика охватывает ядро сперматиды — буду-
Iщую головку спермия.
Всредней зоне акробласта из мельчайших зерен формируется
ак р о с о м а . Это уплотненное тельце, богатое ферментом гиалуронидавой. К противоположному от ядра полюсу перемещается центросома, в которой различают проксимальную и дистальную
£2
*Рис'. 39. Период формирования (схема). Дифференцировка сперматиды и
•образование спермия (^4—Ж).
1 _ ядтзо сперматиды; г — комплекс Гольджи; 8 — центриоли; 4 — зачаток акросомьг б — митохондрии; 6 — жгутик; 7 — акросомный чехлик; 8 — проксималь ная центриоль; 9 — дистальная центриоль; Ю — ядерное вещество, сконцентриро ванное в головне спермия; 11 — митохондриальная спираль; 12 — остатки цито плазмы; 13 — головка; U — шейка; 15 — средняя часть; 16 — хвостик; П — кон цевой участок хвостика.
ц е н т р и о л и . Они образуют шейку спермия. Дистальная цент риоль делится на две части — переднюю и заднюю. От передней части дистальной центриоли отрастает осевая нить хвостика, со стоящая из микротрубочек. Задняя часть дистальной центриоли приобретает вид колечка. Она, сползая по осевой нити хвостика
иувлекая за собой цитоплазму сперматиды, содержащую мито хондрии и гликоген, располагается на границе между начальным
иглавным отделами хвостика спермия. Смещаясь по хвостику, цитоплазма тонким слоем одевает главную его часть. Клетка про должает удлиняться и приобретает форму спермия.
Ра з в и т и е я й ц е к л е т о к , или овогенез (см. рис. 38—//), — -сложный и очень длительный процесс. Он начинается в период эмбриогенеза и завершается в органах половой системы полово зрелой самки. Слагается овогенез из трех периодов: размножения, роста, созревания.
Период размножения протекает в период внутриутробного раз вития и завершается в течение первых месяцев после рождения. Клетки называются овогониями (оогониями). Они незначитель-
53
ЕЫХ размеров, бедны цитоплазмой, их ядра имеют диплоидное число хромосом. Оогонии интенсивно митотически делятся. Неко торые из них перестают делиться, покрываются клетками фолли кулярного эпителия, которые снабжают яйцеклетку питательным материалом.
В период роста клетки называются п е р в и ч н ы м и ооцитами. Они увеличиваются в размере. Изменения в ядрах анало гичны первичным сперматоцитам; они проходят также лептотенную, зиготенную, пахитенную и диплотенную стадии. Затем в ооците начинается интенсивный синтез и накопление желтка, необходимого для течения первых стадий развития зародыша, в связи с чем хромосомы вновь деспирализуются. Этот процесс про
текает в две фазы: стадия малого роста (превителлогенез) |
и ста |
|
дия |
большого роста (вителлогенез). Морфологические особенно |
|
сти |
п р е в и т е л л о г е н е з а — локализация органелл |
вокруг |
ядра, активизация функции белоксинтезирующей системы цито плазмы: развиваются рибосомы, гранулярная эндоплазматическая сеть, поэтому цитоплазма окрашивается основными красителями, то есть она базофильна. Хорошо развиты комплекс Гольджи и ми тохондрии. Ядро округлое, содержит мало хроматина, хорошо развито ядрышко. Вторичная оболочка ооцита состоит из одного слоя фолликулярных клеток. Превителлогенез обычно длится до наступления половой зрелости самки.
В и т е л л о г е н е з — стадия интенсивного накопления желтка в цитоплазме первичного ооцита. В данном процессе принимает участие весь организм. Фолликулярные клетки в ооцит транспор тируют желток. Они интенсивно делятся митозом, поэтому вто ричная оболочка становится многослойной.
Органеллы из перинуклеарной зоны перемещаются на пери ферию: формируется кортикальный слой, который играет важную роль в начальных стадиях эмбриогенеза.
Период созревания состоит из двух быстро следующих друг за другом делений созревания, в ходе которых диплоидная клетка становится гаплоидной. Этот процесс обычно протекает в яйцеводе после выхода первичного ооцита из яичника.
Первое деление созревания завершается образованием двух неравноценных структур — вторичного ооцита и первого напра вительного, или редукционного, тельца. Каждая из них по срав нению с первичным ооцитом содержит половину количества ядер ного вещества. За первым делением очень скоро наступает вто рое, в ходе которого также образуются только одна зрелая яйце клетка и второе неправильное тельце. Первое направительное тельце к этому моменту делится еще один раз и образуются два направительных тельца.
Следовательно, из одного первичного ооцита в процессе созре вания возникают только одна зрелая яйцеклетка и три направи тельных тельца.
Таким образом, овогенез отличается от сперматогенеза рядом особенностей: начинается в период внутриутробного развития
«ам-ки, имеет длительный период роста, в процессе которого на капливается питательный материал (желток); протекает в яични ке эмбриона и половозрелого животного, а затем в яйцеводе; из одного первичного ооцита образуется только одна зрелая яйце клетка; все яйцеклетки генетически однородны, так как имеют Х-хромосому; женская половая клетка не способна к самостоя тельному передвижению.
РАННИЕ ПЕРИОДЫ ЭМБРИОНАЛЬНОГО РАЗВИТИЯ (ЭМБРИОГЕНЕЗ)
ОПЛОДОТВОРЕНИЕ
Оплодотворение — слияние половых гамет (спермия и яйцеклет ки) и образование нового одноклеточного организма (зиготы) или оплодотворенной яйцеклетки (рис. 40). От зрелой яйцеклетки она отличается удвоенной массой ДНК, диплоидным числом хромо сом, высокой метаболической активностью. У многих животных оплодотворение яйцеклетки внутреннее, и оно протекает в яйце воде при пассивном ее продвижении по направлению матки. Дви жение спермиев в половых путях самки осуществляется благо даря функции аппарата движения этой клетки, положительного хемотаксиса (движение по направлению химических раздражи телей) , реотаксиса (движение против тока жидкости), перисталь тическим сокращениям стенки матки; движению ресничек кле ток, покрывающих внутреннюю поверхность яйцевода.
При сближении половых клеток ферменты акросомы головки спермия разрушают вторичную оболочку яйцеклетки. В момент прикосновения спермия к плазмолемме яйцеклетки на ее поверх ности образуется выпячивание цитоплазмы — воспринимающий бугорок, или бугорок опло дотворения. В ооцит проникают головка, шейка, несущая центросому, и начальная часть хвостового отдела. У млекопитающих в оплодотворении участвует только один спермий. Этот процесс носит название моно- с п е р м и и (рис. 41). Как уже говорилось выше, яйцеклетки всегда содержат только Х-половую хромосому, а спермин либо Х-по-
ловую хромосому, либо Y-половую хромосо му. Если в яйцеклетку проникает спермий с Y-половой хромосомой, то зигота будет со держать Х,У-половые хромосомы, и разви вается самец. При наличии в зиготе Х,Х-по- ловых хромосом будет развиваться самка.
Рис. 40. Оплодотворение яйцеклетки спермием (схема).
54
У птиц, |
рептилий, хвостатых амфибий наблюдается п о л и |
с п е р м и я : |
в яйцеклетку проникает одновременно несколько |
спермиев. У птиц все спермин имеют Z-хромосому, а яйцеклеткиили Z-доловую хромосому, или W-хромосому. Таким образом, но вый организм получает хромосомы мужской и женской половых клеток и поэтому наследует признаки обоих родителей.
После проникновения спермия в яйцеклетку вокруг последней формируется оболочка оплодотворения, препятствующая проник новению в ооцит других спермий. Яйцеклетки выделяют особые вещества — агглютинины. Они склеивают другие спермин, а фол ликулярные клетки вторичной оболочки яйцеклетки их погло щают.
Рис. 41. Спермий, втягиваемый в цитоплазму яйцеклетки воспринимающим бугорком (электронная микрофотография):
1 — воспринимающий бугорок; г — центриоль спермия; 3 — оболочка оплодотво рения; 4 — митохондриалыюе тельце; 5 — ядро спермия; б — цитоплазма яйце* клетки (по Вилли и Детье).
1 — цитоплазма-яйцеклетки; 2 — ядро яйцеклетки; з — блестящая оболочка; 4 — фолликулярный эпителий; 5 — головка спермия; в —шейка спермия; 7 — хвост спермия; 8 — воспринимающий бугорок; 9 — оболочка оплодотворения; 10 — жен ский и 11 — мужской пронуклеусы; 12 — веретено между центриолями; 13 — синкарион (по Елисееву).
Головка спермия, внедрившаяся в цитоплазму яйцеклетки, на бухает, округляется и приобретает форму округлого ядра — муж ской пронуклеус (рис. 42). Последний перемещается к округлому ядру яйцеклетки — женскому пронуклеусу. Оба пронуклеуса гаплоидны, и после их соединения образуется диплоидное ядро зиготы. Процесс соединения пронуклеусов называется с и н к а- ри-онои. В это время в клетке резко усиливаются метаболиче ские процессы.
Яйцеклетка лишена центриолей. В нее вносят их спермий. Центриоли шейки спермия расходятся, образуется ахроматиновое веретено; ядерная мембрана исчезает, отцовские и материнские хромосомы формируют материнскую звезду первого митотического деления оплодотворенной яйцеклетки. Так начинается следующий этап эмбриогенеза — дробление.
При полиспермии (рептилии, птицы и др.) в яйцеклетку про никает одновременно несколько спермиев, однако с ее ядром соединяется только один из них. Неоплодотворенные яйцеклетки и неучаствующие в оплодотворении спермий гибнут и в яйцеводах или матке поглощаются клетками крови—лейкоцитами (leukos— белый, kytos — клетка).
Таким образом, при оплодотворении осуществляется: 1) акти вация яйцеклетки, после чего начинается ее деление (дробление);
57
5 С
2) рекомбинация отцовских и материнских хромосом, что в ко нечном результате обусловливает адаптацию особи к изменениям окружающей среды и их выживаемость. Таково обще биологиче ское значение оплодотворения.
ДРОБЛЕНИЕ
Дробление — дальнейший процесс развития одноклеточной зиго ты, в ходе которого образуется многоклеточная бластула, которая состоит из стенки — б л а с т о д е р м ы и полости — б л а с т о- ц е л я. В бластодерме различают крышу, дно и расположенную между ними краевую зону. В процессе митотического деления 8иготы образуются новые клетки — бластомеры, остающиеся тес но связанными друг с другом.
В начальной стадии дробления многоклеточный организм посвоему размеру сходен с зиготой, так как его бластомеры, делясь,, не достигают размера исходной клетки. Характер дробления в эволюционном ряду хордовых различен, что в значительной сте пени обусловлено количеством и распределением желтка в яйце клетках.
Дробление может быть полным (голобластическим) или час тичным (меробластическим). При голобластическом дроблении принимает участие весь материал зиготы, при меробластическом— только та ее зона, которая лишена желтка.
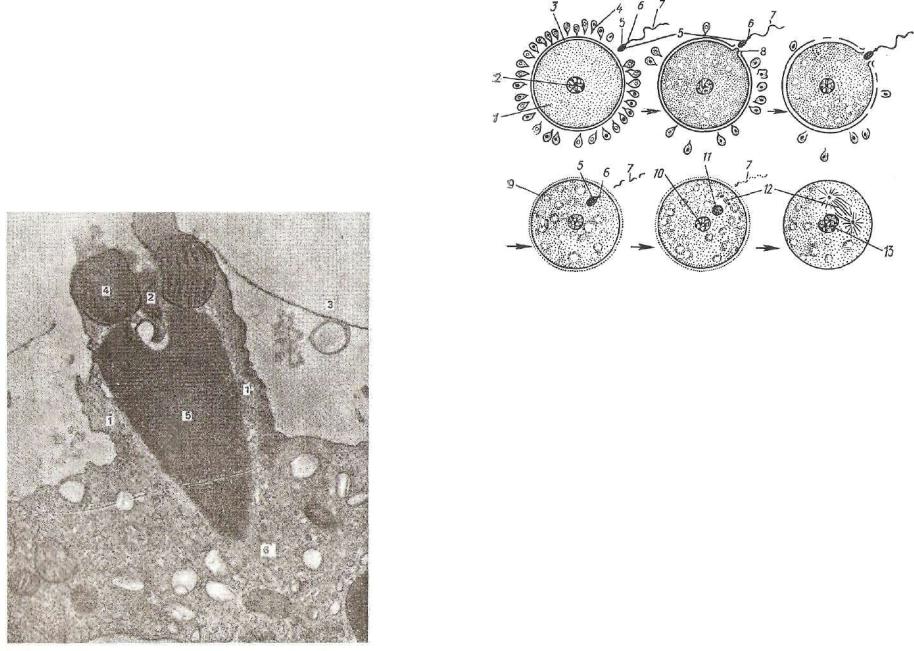
Полное дробление классифицируют на равномерное и неравно мерное. П о л н о е р а в н о м е р н о е д р о б л е н и е (рис. 43) характерно для яиц с малым количеством желтка (олиголецитальных) и равномерно расположенным по всей цитоплазме клетки желтком (изолецитальных). Примером такого дробления могут служить ланцетник, аскарида и др. В оплодотворенной яйцеклет ке различают два полюса: верхний — анимальный и нижний — вегетативный.
После оплодотворения желток, незначительное количество которого было равномерно распределено по всей цитоплазме, перемещается к вегетативному полюсу. Первая борозда дробления проходит в меридиональном направлении и делит зиготу на два бластомера, которые соответствуют будущей левой и правой поло вине тела зародыша. Вторая борозда дробления проходит также меридионально под прямым углом к первой, и теперь зародыш состоит из четырех бластомеров. Третья борозда дробления имеет экваториальное направление, поэтому каждый бластомер делится на две части. Такой зародыш построен из восьми бластомеров, при этом четыре из них образовались из вегетативного полюса зиготы, в связи с чем они содержат весь желток зиготы и отличаются большими размерами. Эти бластомеры соответствуют задней час ти тела; аиимальиые — четыре — передней части.
Затем появляются две меридиональные борозды, делящие заро дыш на 16 бластомеров. Пятое дробление — это две широтные борозды, в составе зародыша 32 бластомера. Они начинают посте-
Рис. 43. Схема расположения борозд дробле ния у ланцетника (А):
1 — зародыш на стадии двух бластомеров; XI — зародыш на стадии четырех бластомеров; 1X1 — зародыш на стадии восьми бластомеров; IV — вародыш на стадии 16 бластомеров; У — зародыш на стадии 32 бластомеров; VI — зародыш на ста дии 64 бластомеров; VII — зародыш на стадии 128 бластомеров. Строение бластулы (В): 1 — бла стодерма; г — бластоцель; 3 — дно; 4 — краевая зона; 5 — крыша бластулы.
пенно отодвигаться друг от друга, контактируя лишь боковыми поверхностями. Внутри зародыша образуется сначала небольшая полость — бластоцель, которая постепенно увеличивается. После шестого дробления образуется 64 клетки, при этом борозды дроб ления проходят меридионально. После седьмого дробления (воз никают четыре широтные борозды) зародыш состоит из 128 блас томеров.
Позднее синхронность в делении зародыша нарушается, блас томеры отодвигаются на периферию и располагаются в один слой, формируя бластодерму, а в центре зародыша образуется бласто цель.
Дробление завершается образованием бластулы, форма кото рой напоминает шар, заполненный жидкостью. Стенка шара обра зована клетками бластодермы.
Таким образом, при полном равномерном дроблении материал всей зиготы участвует в делении и после каждого деления (дроб ления) число клеток (бластомеров) увеличивается вдвое.
В бластодерме дифференцируются следующие участки: кры ша, построенная из относительно мелких бластомеров; дно — это более крупные бластомеры и краевая зона, лежащая между дном и крышей бластулы.
58 |
59 |
П о л н о е н е р а в н о м е р н о е д р о б л е н и е |
характерно |
для мезолецитальных (среднее количество желтка) |
и телолеци- |
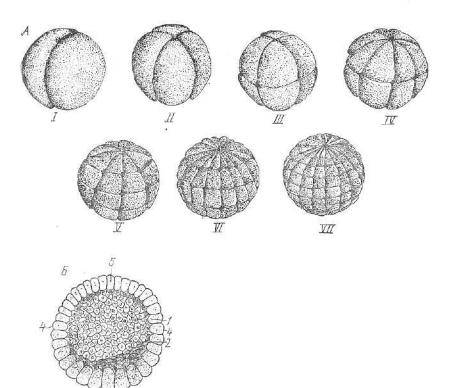
тальных (желток расположен в вегетативном полюсе) яйцекле ток. Примером этого типа дробления может служить дробление зиготы амфибий (рис. 44).
Дробление начинается с образования двух меридиональных борозд дробления, следующих друг за другом под прямым углом. Они быстро делят лишенный желтка анимальный полюс зиготы на два, а затем па четыре мелких бластомера. Вегетативный полюс, содержавший весь желток зиготы, дробится значительно медленнее, и бластомеры, возникающие здесь, более крупных размеров.
Третья борозда проходит ближе к анимальному полюсу зиго ты и имеет широтное направление. Широтные борозды дробления сменяются меридиональными, при этом очень скоро возникает асинхронность и тангенциальность (деление бластомеров в плос-
60
Рис. 45. Частичное (дискондальное) дробяеняе зароды ша курицы:
А, В — стадии дробления — вид сверху (А — две мери диональные борозды, В — бо лее поздняя стадия дробле ния); С — разрез зародыше вого диска (а, Ь, с, — крае вые клетки, расположенные на желтке; d, e, f, g, h — клетки, изолированные от желтка).
кости, параллельной по верхности зиготы) в дроблении, поэтому оно завершается образовани ем многослойной бласту
лы. Крыша бластулы построена из мелких бластомеров, именуе мых м и к р о м е р а м и . Дно состоит из крупных бластомеров — м а к р о м е р о в . Весь желток локализован в макромерах. Бластоцель сдвинут к анимальному полюсу и уменьшен в размере. Бла стула, образовавшаяся в процессе голобластического (полного) дробления, носит название ц е л о б л а с т у л ы .
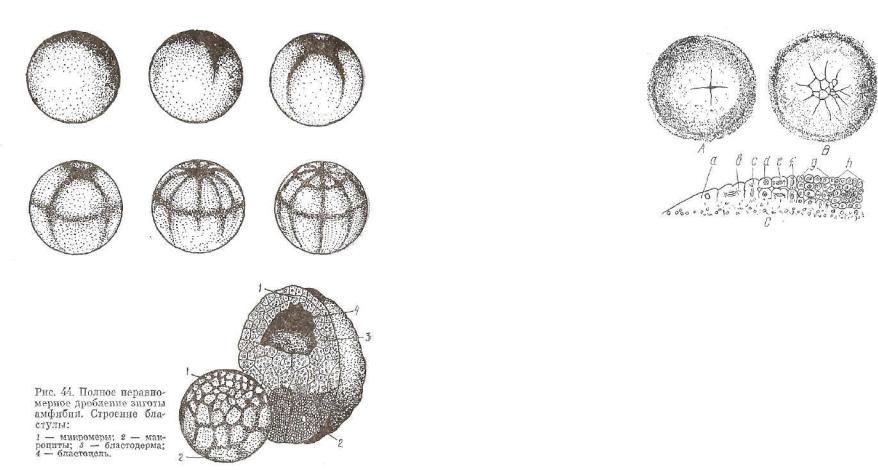
Частичное, или меробластическое (дискоидальное), дробление*- распространено у рыб, рептилий, птиц и характерно для полилеЧитальных (много желтка) и телолецитальных яиц (рис. 45).
В дроблении участвует только лишенный желтка поверхност ный слой анимального полюса зиготы, так как здесь находятся ядро клетки и цитоплазма без желтка. Вся остальная часть зиго ты загружена желтком и поэтому не дробится.
Первые две меридиональные борозды проходят через анималь ный полюс под углом один к другому. Они не распространяются на вегетативный полюс, в связи с чем последний остается нераз деленным на бластомеры. Меридиональные борозды сменяются широтными и тангенциальными. Бластомеры, образовавшиеся в- ходе дробления, располагаются на желтке в один слой. Этот слой называется зародышевым диском, поэтому дробление получил» название д и с к о и д а л ь н о г о .
На построение тела зародыша используется только его цен-
'тральная часть — зародышевый щиток. Остальная часть зароды шевого диска участвует в образовании временных (провизорные) органов — зародышевых оболочек, которые создают благоприят ные условия для развития зародыша.
Дробление завершается образованием бластулы, у которой1 бластоцель имеет вид узкой щели и сдвинут к анимальному по люсу. Крыша бластулы построена из бластомеров. Краевая зо на — это интенсивно делящиеся клетки (бластомеры) перифери ческой зоны зародышевого диска. Дном является неразделенный' на бластомеры желток вегетативного полюса зиготы. Такой т и т бластулы называется д и с к о б л а с т у л о й .
6*