

gistologia
.pdfслой. Последний в комплексе с базальной мембраной и эндотелиальными клетками стенки капилляров входит в состав гематотимусного барьера, препятствующего проникновению анти генов в пространства коркового вещества, где происходят раз множение и дифференцировка Т-лимфоцитов. Из лимфоидных клеток наиболее крупные — лимфобласты, располагаются в самой наружной зоне коркового вещества. Показано, что они образу ются из проникающих сюда предшественников Т-лимфоцитов костно-мозгового происхождения. Под влиянием гуморальных факторов (тимозин и др.), выделяемых эпителиальными клетка ми, в этой зоне происходит антигеннезависимая пролиферация активированных лимфоцитов и превращение их в иммунокомпетентные Т-лимфоциты. Корковое вещество долек тимуса являет ся зоной с наиболее высокой скоростью обновления малых лим фоцитов. Однако подавляющее большинство новообразованных лимфоцитов погибает в пределах этого органа, продукты их рас пада утилизируются макрофагами. Считают, что таким образом разрушаются Т-лимфоциты, запрограммированные на взаимодей ствие с макромолекулами (антигенами) собственного организма. При попадании этих Т-лимфоцитов в кровоток развивается ауто иммунная реакция.
Небольшое количество (до 5%) Т-лимфоцитов, имеющих в плазмолемме рецепторы к чужеродным антигенам, мигрирует из внутренней зоны коркового вещества в кровоток. Циркулируя в крови, они попадают во вторичные органы иммунной системы (селезенку, лимфатические узлы), где заселяют тимусзависимые зоны и в соответствии с поверхностными маркерами превраща ются в подклассы: киллеры, хелперы, супрессоры.
Лимфоциты мозгового вещества обладают очень низкой митотической активностью и относятся к рециркулирующей популя ции Т-лимфоцитов. Характерные образования для мозгового ве щества долек тимуса — тимусные тельца (corpusculum thymicum) — тельца Гассаля, состоящие из концентрически наслоенных друг на друга уплощенных эпителиальных клеток. Перифериче ские живые клетки тимусного тельца имеют светлые ядра и слабооксифильную цитоплазму, в которой гистохимическими мето дами выявляются гликозаминогликаны. В клетках центральной части крупного тельца наблюдаются дистрофические изменения, сопровождающиеся исчезновением ядер и образованием гомоген ной оксифильной массы. Величина и строение отдельных тимусных телец сильно варьируют. Таким образом, корковое и мозго вое вещество долек тимуса отличается составом и структурными особенностями эпителиальной основы и биологическими свойст вами свободных лимфоидных клеток.
Тимус относится к органам, величина которых значительно изменяется с возрастом. Масса тимуса увеличивается в ранний постэмбриональный период до полового созревания — у северных оленей с 15,5 до 55 г (И. С. Решетников); у мышей с 10 до 70 мг. После этого наблюдается прогрессирующее уменьшение долек
органа — в о з р а с т н а я и н в о л ю ц и я . |
В гистологическом от |
||
ношении изменения |
происходят преимущественно |
в корковом |
|
веществе долек, в котором значительно |
уменьшается |
количество- |
|
лимфоцитов. Дольки |
выглядят сморщенными, в них |
обнаружи |
|
вают эпителиальные |
клетки и тимусные тельца, а также тучные |
||
клетки и макрофаги с вакуолизированной |
цитоплазмой. Соедини |
||
тельнотканные междольковые прослойки становятся более во локнистыми и в них увеличивается число жировых клеток. Про должительность возрастной инволюции варьирует в зависимостиот вида животных.
Под влиянием различных внешних и внутренних сильных воздействий (тяжелые травмы, облучения, интоксикации, голо дание, острые инфекционные болезни, сезонные изменения, рез кое увеличение в крови глюкокортикоидных гормонов и др.) мо жет произойти быстрая а к ц и д е н т а л ь н а я и н в о л ю ц и я » тимуса, связанная с интенсивной миграцией лимфоцитов из кор ковой части долек в кровь и их массовой гибелью в самом органе. Нередко акцидентальная инволюция является обратимым про цессом.
КЛОАКАЛЬНАЯ (ФАБРИЦИЕВА) СУМКА ПТИЦ
Клоакальная сумка представляет характерный для птиц лимфо-
эпителиальный |
орган, имеющий |
форму карманообразного выпя |
||||||
чивания, расположенного в дорсальной части |
стенки клоаки. По |
|||||||
строению |
это |
полостной |
орган, |
слизистая |
оболочка |
которого |
||
имеет первичные и вторичные складки, выступающие в |
просвет |
|||||||
его. В складках, покрытых многорядным |
призматическим эпите |
|||||||
лием, находятся тесно |
прилегающие друг |
к |
другу многочислен |
|||||
ные лимфатические узелки, состоящие из |
коркового и мозгового- |
|||||||
вещества |
(рис. |
208). В |
более светлом мозговом веществе обна |
|||||
руживают |
удлиненные |
или |
отростчатые |
эпителиальные |
клетки, |
|||
между которыми располагаются крупные и средние лимфоциты.. В корковом веществе высокая концент рация малых лимфоцитов.
При развитии сумки вначале форми руется эпителиальная основа долок, позднее заселяющаяся мигрирующими из желточного мешка стволовыми клет ками, из которых развиваются многочис ленные лимфоциты.
Фабрициева сумка — центральный орган иммунологической защиты. В ней
Рис. 208. Разрез складки клоакальной фабрицисвой сумки курицы:
1 — лимфатические узелки о корковым и мозговым веществом; 2 — эпителий.
284 |
285^ |
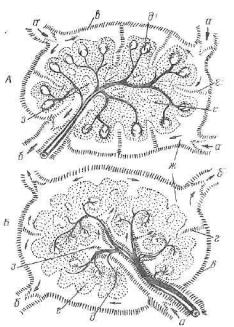
развиваются иммуноциты гуморального иммунитета — бурсо- -зависимые лимфоциты (В-лимфоциты), способные под дейст вием антигенов во вторичных лимфоидных органах превращаться
..в антителообразующие плазматические клетки.
ЛИМФАТИЧЕСКИЕ УЗЛЫ
У позвоночных лимфатические узлы впервые появляются у птиц. "У некоторых водоплавающих (чирки, утки) существуют две пары
.лимфоузлов — шейные и поясничные вытянутой формы и распо-
.ложены по ходу лимфатических сосудов. У кур образования типа -лимфатических узлов отсутствуют, но имеются многочисленные -лимфоидыые скопления, .расположенные в соединительнотканных прослойках печени, легких, почек, яичника, слизистой кишечника.
Лимфатические узлы млекопитающих — многочисленные хо рошо оформленные органы, расположенные в местах слияния <жрупных лимфатических сосудов. Число и размеры лимфоузлов весьма вариабельны у различных видов животных и в разных топографических областях организма. В зависимости от локали зации различают поверхностные и глубокие лимфоузлы. Форма •узлов чаще овальная или бобовидная, брыжеечные лимфоузлы имеют удлиненную форму.
Располагаясь на пути тока лимфы, лимфоузлы являются важ нейшими барьерно-фильтрационными органами, в которых за держиваются и подвергаются фагоцитозу микроорганизмы, чуже- «родные частицы, разрушающиеся клетки.
Лимфатические узлы — периферические органы лимфоидного кроветворения и иммунной защиты. В ответ на антигенное воз действие в лимфоидной ткани узла образуются иммуноциты гуморального иммунитета .(плазматические клетки) и система эффекторных Т-кяеток (киллеров, хелперов, супрессоров).
Развитие всего комплекса лимфоузлов у животных растяги вается на значительный промежуток времени, но большинство
.лимфоузлов формируется во второй половине эмбрионального периода. Образование закладки лимфоузла связано с размноже нием и уплотнением клеток мезенхимы вокруг лимфатических ^сосудов. В дальнейшем лимфатические сосуды на периферии за кладки сливаются и образуют краевой синус, а стенки их вместе •с окружающей мезенхимой формируют капсулу и отходящие от
.нее перекладины — трабекулы.
Сеть лимфатических |
сосудов внутри развивающегося |
узла |
|||
'представляет собой систему |
промежуточных |
синусов, которые |
|||
•разделяют образовавшуюся |
из мезенхимы ретикулярную |
ткань |
|||
на овальные образования |
в |
периферической |
части |
закладки и |
|
мозговые тяжи в ее центральной части. Позднее эти |
структурные |
||||
• образования заселяются кроветворными клетками. В-лимфоциты вселяются преимущественно в самую поверхностную и централь ную зону узла, а Т-лимфоциты — в промежуточную между ними •область (будущая паракортикальная зона). Дальнейший период
Рис. 209. Схема строения лим фатического узла:
А — собаки; В — свиньи; а — приносящие лимфатические сосу ды; б — выносящий лимфатиче ский сосуд; в — капсула; г — трабекулы; д — лимфатические фолликулы; е — мякотные тяжи; ж — краевой лимфатический си нус; з — воротный синус.
развития характеризуется главным образом количест венными изменениями. По степенно увеличивается число лимфоидных клеток и макрофагов. В корковом веществе формируется ос нова фолликулов, в кото рых видны клетки со свет лым ядром — дендритные макрофаги.
Микроскопическое стро ение лимфоузла. Лимфа из определенных областей ор ганизма поступает в лим
фоузел по приносящим лимфатическим сосудам, впадающим навыпуклой стороне узла. У свиней сосуды, приносящие лимфу,., впадают в вогнутую часть — ворота узла, а сосуды, выносящиелимфу, выходят на противоположной выпуклой стороне. В связл* с этим общая микроскопическая структура лимфоузлов у свиней" отличается от структуры узлов других видов животных (рис. 209).
Лимфатический узел характеризуется наличием следующих- структурно-функциональных компонентов: а) соединительно тканного остова, состоящего из капсулы и системы трабекул,. разграничивающих внутреннюю полость узла на сообщающиеся1 между собой отсеки; б) лимфоидной ткани, из которой формиру ются характерные структурные образования в периферической' промежуточной и центральной части органа; в) системы лимфа тических синусов, обеспечивающих условия нормального тока, лимфы, необходимого для функционирования лимфоузла.
К а п с у л а , окружая узел со всех сторон, образует утолщение • в области ворот. У различных видов животных степень развития'
ее неодинакова, |
в частности у крупного рогатого скота она |
отли |
|||
чается значительной толщиной. |
Более толстая капсула в поверх |
||||
ностных узлах. |
От внутренней поверхности капсулы |
в |
глубь- |
||
узла отходят перегородки — т р а б е к у л ы . |
Различают |
трабеку |
|||
лы хиларные, отходящие от соединительной |
ткани ворот |
узла,. |
|||
и капсулярные. |
Хиларные более |
массивные, содержат кровенос |
|||
ные сосуды и глубоко проникают во внутреннюю часть узла,, капсулярные обычно тонкие и неглубоко проникают в корковую-
287"
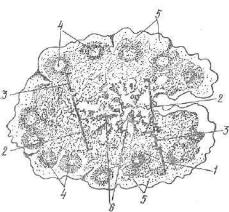
Рис. 210. Схема расположе ния структурно-функциональ ных зон и образований лимфа тического узла:
1 — корковое вещество; 2 — моз говое вещество; 3 — паракортикальная зона; 4 — светлые цент ры лимфатических фолликулов; 5 — подкапсулярный лимфатиче ский синус; б — мозговые тяжи (рис. Козлова).
часть узла. Капсула и трабекулы состоят из плотной соединительной ткани, в которой преобладают коллагеновые волокна, между
волокнами располагаются фиброциты. Во внутренней зоне капсу лы развита сеть эластических волокон, переходящая в эластиче ские элементы приносящих и выносящих лимфатических сосудов, а также имеются пучки гладкомышечных клеток, сокращение ко торых способствует продвижению лимфы.
Л и м ф о и д н а я т к а н ь узла состоит из многочисленных клеток лимфоидного ряда, макрофагов в комплексе с ретикуляр ными клетками, сетью ретикулярных волокон и своеобразно ор ганизованной капиллярно-сосудистой сетью. В наружной зоне из лимфоидной ткани построены специализированные округлые об
разования — лимфатические фолликулы |
(folliculus |
thymphaticus), |
|||||||
в центральной зоне — анастомозирующие |
между собой |
мозговые |
|||||||
•тяжи (chorda medullaris). Область расположения |
лимфатических |
||||||||
фолликулов в комплексе с другими |
структурными образованиями |
||||||||
наружной |
зоны |
узла называется |
к о р к о в ы м |
в е щ е с т в о м |
|||||
(cortex), а зона |
мозговых тяжей в комплексе |
с |
другими струк |
||||||
турными |
образованиями |
центральной части |
узла — м о з г о в ы м |
||||||
в е щ е с т в о м |
(medulla). Расположенная между корковым и моз |
||||||||
говым веществом зона диффузной лимфоидной ткани |
именуется |
||||||||
п а р а к о р т и к а л ь н о й |
з о н о й (рис. 210). Соотношения меж |
||||||||
ду размерами |
коркового вещества, паракортикальной зоны и моз |
||||||||
гового вещества в различных узлах |
одного и |
того же |
животного |
||||||
и одноименных узлах разных животных неодинаковы. |
Имеются |
||||||||
также выраженные возрастные различия в |
соотношении этих |
||||||||
структурных |
частей. |
|
|
|
|
|
|
||
• Лимфатические фолликулы в корковом веществе вследствие неравномерного распределения разных типов клеток имеют на окрашенных препаратах вид неоднородных образований. Разли чают лимфатические фолликулы без светлых центров, состоящие в основном из лимфоидных клеток, и лимфатические фолликулы со светлым центром (термин «светлый центр» заменил прежние названия: «зародышевый центр», «реактивный центр», «центр размножения», «герминативный центр»). Развитие светлых цент-
ров происходит под влиянием антигенного воздействия на орга низм. Они отсутствуют в лимфатических фолликулах в эмбрио нальный период и не обнаруживаются в постэмбриональный пе риод у животных, выращенных в стерильных условиях. Структура фолликулов поддерживается с помощью сети ретикулярных во локон, количество которых наибольшее в его периферической части. Для центра фолликулов характерно наличие особых ден дритных клеток сложной отростчатой формы. Такие клетки свой ственны лишь В-зонам вторичных лимфоидных органов. С по мощью рецепторов клеточной оболочки дендритные клетки могут фиксировать антиген и длительное время прочно удерживать об разующийся комплекс антиген — антитело, создавая условия для эффективного контакта его с В-лимфоцитами. В светлом центре содержатся свободные макрофаги, способные к фагоцитозу про дуктов распада лимфоцитов, погибающих при массивном анти генном раздражении. Цитоплазма таких макрофагов вакуолизирована и содержит плотные окрашенные тельца. Здесь же нахо дятся бластные клетки, имеющие крупные размеры и заметный ободок базофильной (пиронинофильной) цитоплазмы. Некоторые из этих клеток находятся в различных стадиях митотического деления. В периферической (мантийной) зоне фолликулов распо лагаются преимущественно малые лимфоциты, относящиеся к В-клеткам. Таким образом, фолликулы лимфоузла относятся к тимуснезависимым образованиям лимфоидной системы. Микро структура лимфатического фолликула очень динамична и зави сит от функционального состояния лимфоузла, что, в свою оче редь, определяется фазой иммунного ответа на антигены.
Паракортикальная зона относится к тимусзависимой и запол нена Т-лимфоцитами, которые принадлежат к рециркулирующему фонду клеток. В этой зоне располагаются особые разновидно сти макрофагов — интердигитирующие клетки, имеющие много численные пальцевидные отростки. Ядра этих клеток светлые с краевым расположением хроматина. Полагают, что своими гумо ральными факторами (гликопротеиды) эти клетки индуцируют пролиферацию и дифференциацию Т-лимфоцитов в эффекторные клетки (киллеры) клеточного иммунитета. Для паракортикальной зоны характерны сосуды особого типа — посткапиллярные венулы, выстланные кубическим эндотелием, через клетки которого рециркулирующие лимфоциты выходят из крови в лимфоидную ткань узла. Структура этой зоны поддерживается хорошо разви той сетью ретикулярных волокон. Размеры и клеточный состав паракортикальной зоны непостоянны и зависят от стадии и ха рактера иммунного ответа. При формировании реакций клеточ ного иммунитета в этой зоне происходят стимуляция Т-лимфо- цитов, их превращение в бластные формы, пролиферация и обра зование эффекторных клеток.
Мозговые тяжи представляют собой лентовидные, анастомо зирующие образования из лимфоидной ткани, расположенные по ходу мелких кровеносных сосудов в мозговом веществе. Форма
388 |
19 Заказ № 908 |
289 |
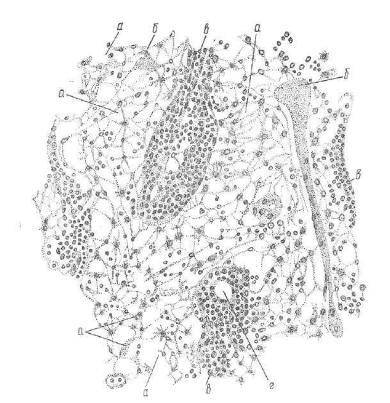
Рис. 211. Участок мозгового вещества лимфатического узла собаки (по Мак симову) :
а — ретикулярная ткань в промежуточных синусах; б — трабенулы; в — мякотныев (мозговые) тяжи; г — артерия в составе мякотного тяжа.
тяжей поддерживается с помощью обильного сплетения ретику лярных волокон, которые отделяют тяжи от синусов мозгового вещества. В петлях ретикулярной сети располагаются многочис ленные лимфоциты (В-лимфоциты), макрофаги и плазматические клетки. По ходу тяжей от коркового вещества к воротам завер шается образование зрелых плазматических клеток, поэтому моз говые тяжи являются основной зоной их локализации в лимфо узле.
Циркуляция лимфы в лимфатическом узле является одним из главных факторов функционирования этого органа (рис. 211). Пути тока лимфы в узле называются лимфатическими синусами.
Различают |
к р а е в о й с и н у с — пространство между капсулой и |
корковыми |
фолликулами, его пересекают тра-.бекулы, проникаю |
щие в узел; п р о м е ж у т о ч н ы е |
к о р к о в ы е с и н у с ы — |
между трабекулами и фолликулами; |
п р о м е ж у т о ч н ы е моз - |
290 |
|
«г о в ы е с и н у с ы — между мозговыми тяжами и трабекулами и в о р о т н ы й с и н у с , из которого лимфа поступает в выносящие лимфатические сосуды. Суммарный диаметр всех синусов узла значительно превосходит диаметр приносящих сосудов, что обес печивает замедленное внутриузловое течение лимфы и возмож ность длительного ее контакта с различными клетками. Стенки синусов выстланы уплощенными береговыми клетками, с уча стием которых на наружной стороне краевого синуса формирз'- •ется сплошной клеточный слой, расположенный на базальной мембране. На внутренней поверхности краевого синуса и на поверхностях промежуточных синусов, обращенных к фоллику лам и мозговым тяжам, базальная мембрана отсутствует и слой береговых клеток прерывистый. Через узкие промежутки между береговыми клетками осуществляется выход лимфоцитов из фол ликулов, паракортикальной зоны и мозговых тяжей в полость си нусов. В просвете синусов располагаются фиксированные отростчатые клетки, соединенные со стенками, а также свободные мак рофаги, лимфоциты и другие клетки. Благодаря наличию много численных макрофагов в синусах задерживается и поглощается большая часть попадающих в лимфоузел антигенов.
Кровоснабжению лимфатического узла свойственны некото рые особенности. Артерии, проникая в ворота, разветвляются на множество веточек, проходящих по хиларным трабекулам в раз личные зоны узла. Ветви, направляющиеся к мозговому вещест ву, короткие и сразу же распадаются на капилляры мозговых тяжей. Ветви коркового вещества вначале не ветвятся и идут к фолликулам. В каждый фолликул вступают 1—3 артериолы, ко торые распадаются на капилляры, последние достигают светлого центра. Капилляры впадают в посткапиллярные венулы, распо ложенные в поверхностной зоне фолликула, вблизи синусов. Пост капиллярные венулы в паракортикальной зоне проходят на гра нице с мозговым веществом. Чбрез высокий эндотелий венул осуществляется избирательное прохождение лимфоцитов из по лости венул в лимфатическую ткань и далее в синусы. С выпук лой стороны узла проникают артерии через капсулу и развет вляются на капилляры в самой капсуле и капсулярных трабекулах.
ГЕМОЛИМФАТИЧЕСКИЕ УЗЛЫ
У животных (круппого рогатого скота, лошадей, свиней и др.) преимущественно в грудной полости, вдоль грудной и брюшной: аорты, почечных сосудов встречаются небольшие узлы желтого или красного цвета. По развитию и строению они сходны с обыч ными лимфатическими узлами. Снаружи гемолимфатические узлы покрыты соединительнотканной капсулой, содержащей гладкую мышечную ткань. В периферической части узла имеются отдельные фолликулы. Между тонкими мозговыми тяжами рас положены широкие промежуточные синусы, полости которых
29* |
291 |
ЛИМФАТИЧЕСКИЕ ОБРАЗОВАНИЯ ПИЩЕВАРИТЕЛЬНОГО ТРАКТА
В соединительнотканных слоях слизистой оболочки на всем про тяжении пищеварительного тракта имеются скопления лимфоид ных элементов в виде или диффузной лимфондной ткани, или оди ночных фолликулов, или в виде группы фолликулов. Крупные агрегаты лимфоидных фолликулов находятся в слизистой обо лочке на границе ротовой полости и глотки. Они называются
м и н д а л и н а м и . Различают |
по месту |
расположения |
небные, |
||
язычные, глоточные и околонадгортанные |
(только у свиней) мин |
||||
далины. В |
совокупности |
все |
они образуют г л о т о ч н о е |
л и м- |
|
ф о и д н о е |
к о л ь ц о (рис.212). |
|
|
||
В слизистой оболочке |
кишечника комплексы лимфоидных |
||||
фолликулов с межфолликулярной лимфоидной диффузной тканью именуются п е и е р о в ы м и б л я ш к а м и . Они наиболее часто встречаются в подвздошной, слепой и ободочной кишке.
По строению и клеточному составу фолликулы кишечной стенки сходны с фолликулами коркового вещества лимфатических узлов. В них часто образуются крупные светлые центры.
В функциональном отношении в лимфатических образованиях стенки пищеварительного канала происходят опосредованные Т- и В-лимфоцитами иммунологические реакции, связанные с про никновением антигенов со стороны полости соответствующих отделов пищеварительного канала. Считают, что здесь в значи тельном количестве образуются плазматические клетки, выраба тывающие иммуноглобулины типа A (IgA), с помощью которых происходит обезвреживание антигенов. В местах расположения лимфоидных образований лимфоциты в большом количестве миг рируют через эпителий и достигают его поверхности.
СЕЛЕЗЕНКА
Селезенка — непарный орган, расположенный в брюшной поло сти на большой кривизне желудка, у жвачных — на рубце. Фор ма ее варьирует от плоской удлиненной до округлой; у животных разных видов форма и размеры могут быть различными. Цвет селезенки — от интенсивного красно-коричневого до сине-фиоле тового — объясняется большим количеством содержащейся в ней крови.
6
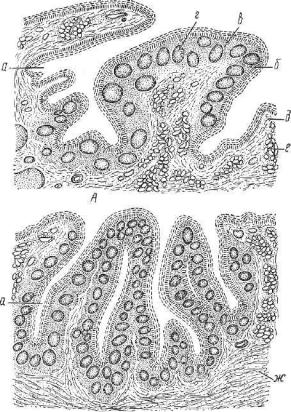
Рис. 212. Нёбные миндалины: |
|
|
|
миндалин; б — |
||||||
А — собаки, |
В — овцы (по |
Элленбергеру и Траутману); а — ямки |
||||||||
эпителий; в |
— ретикулярная ткань; г — лимфатические фолликулы; |
д — рыхлая сое |
||||||||
динительная ткань; е — железы; ж — пучки мышечных волокон. |
|
|
|
|||||||
Селезенка — многофункциональный орган. У большинства жи |
||||||||||
вотных это |
важный орган лимфоцитообразовапия и иммунитета, |
|||||||||
в котором |
|
под влиянием |
антигенов, |
присутствующих |
в |
крови, |
||||
происходит |
образование |
клеток |
либо |
продуцирующих |
гумораль |
|||||
ные |
антитела, либо |
участвующих в реакциях клеточного |
имму |
|||||||
нитета. У |
|
некоторых |
животных |
(грызуны) селезенка — универ |
||||||
сальный |
орган кроветворения, где образуются клетки |
лимфоид- |
||||||||
ного, |
эритроидного и гранулоцитарного ростков. |
Селезенка — |
||||||||
мощный макрофагический орган. При участии многочисленных
макрофагов в |
ней |
происходит |
разрушение клеток |
крови |
и осо |
бенно эритроцитов |
(«кладбище |
эритроцитов»), продукты |
распа |
||
да последних |
(железо, белки) |
вновь используются |
в организме. |
||
292 |
29а |
|
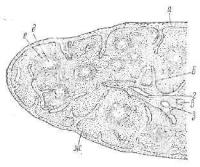
Рис. 213. Селезенка кошки (по Элленбергеру и Траутману):
а — капсула; б — трабекулы; в — трабекулярная артерия; г — трабекулярная вена; д — светлый центр лимфатическо го фолликула; е — центральная арте рия; ж — красная пульпа; а — сосуди стое влагалище.
щ * |
Селезенка — орган депонирова- |
||
§*• |
ния крови. Особенно выражена |
||
|
депонирующая |
функция |
селе- |
С8Я2:Г-*2- |
зенки у лошади и жвачных. |
||
|
Развивается |
селезенка |
пз |
скоплений быстро размножающихся клеток мезенхимы в области
дорсальной |
части брыжейки. |
В |
начальный |
период |
развития |
||||||||
в закладке |
происходит формирование |
из |
|
мезенхимы |
волокни |
||||||||
стого каркаса, |
сосудистого русла |
и |
ретикулярной стромы. Пос |
||||||||||
ледняя |
заселяется стволовыми |
клетками |
и |
|
макрофагами. Вна |
||||||||
чале это орган миелоидного кроветворения. |
Затем |
идет интен |
|||||||||||
сивное вселение из центральных лимфоидных |
органов |
лимфо |
|||||||||||
цитов, |
которые |
сначала |
располагаются |
|
равномерно |
вокруг |
|||||||
центральных артерий (Т-зона). |
В-зоны |
|
образуются |
позднее, |
|||||||||
что связано |
с |
концентрацией макрофагов |
и |
лимфоцитов сбоку |
|||||||||
от Т-зон. |
Одновременно |
с |
развитием |
лимфатических |
узелков |
||||||||
наблюдается |
и |
формирование |
красной |
пульпы |
селезенки. |
||||||||
В ранний постэмбриональный |
период отмечают увеличение коли |
||||||||||||
чества и объема узелков, развитие и расширение в них центров размножения.
Микроскопическое строение селезенки. Основные структурнофункциональные элементы селезенки — опорно-сократительный аппарат, представленный капсулой и системой трабекул, и осталь ная межтрабекулярная часть — пульпа, построенная в основном из ретикулярной ткани. Различают белую и красную пульпу (рис. 213).
Селезенка покрыта серозной оболочкой, плотно срастающей ся с соединительнотканной капсулой. От капсулы внутрь органа отходят перекладины — трабекулы, формирующие своеобразный сетевидный каркас. Наиболее массивные трабекулы у ворот селе зенки, в них расположены крупные кровеносные сосуды — трабекулярные артерии и вены. Последние относятся к венам безмышечпого типа и на препаратах достаточно отчетливо отличаются по строению от стенки артерий.
Капсула и трабекулы состоят из плотной волокнистой соеди нительной и гладкой мышечной ткани. Значительное количество мышечной ткани развивается и содержится в селезенке депони рующего типа (лошадь, жвачные, свиньи, хищные). Сокращение гладкой мышечной ткани способствует выталкиванию депониро ванной крови в кровяное русло. В соединительной ткани капсу лы и трабекул преобладают эластические волокна, позволяющие
селезенке |
изменять |
свои |
размеры |
и выдерживать значительное |
|||
увеличение ее в объеме. |
|
|
|
|
|||
Б е л а я |
п у л ь п а |
(pulpa lienis |
alba) |
макроскопически и на |
|||
неокрашенных |
препаратах |
представляет |
совокупность |
светло-се |
|||
рых округлых |
или овальных образований (узелков), |
незаконо |
|||||
мерно рассредоточенных по всей селезенке. Количество узелков. у разных видов животных различное. В селезенке крупного ро гатого скота их много и они отчетливо отграничены от красной пульпы. Меньше узелков в селезенке лошади и свиньи.
При световой микроскопии каждый лимфатический узелок является образованием, состоящим из комплекса клеток лимфоидной ткани, расположенных в адвентиции артерии и отходящих от нее многочисленных гемокапилляров. Артерия узелка называ ется ц е н т р а л ь н о й , однако чаще она расположена эксцентрич но. В развитом лимфатическом узелке различают несколько струк турно-функциональных зон: периартериальную, светлый центр с мантийной зоной и маргинальную зону. П е р и а р т е р ж а л ь н а я
з о н а — своеобразная |
муфта, состоящая из малых лимфоцитов,, |
|||
тесно прилегающих |
друг к другу |
и |
интердигитирующих |
клеток. |
Лимфоциты этой зоны относятся |
к |
рециркулирующему |
фонду |
|
Т-клеток. Сюда они проникают из гемокапилляров, а после анти генной стимуляции могут мигрировать в синусы красной пульпы, Интердигитирующие клетки — особые отростчатые макрофаги, поглощающие антиген и стимулирующие блаеттрансформацию, пролиферацию и превращение Т-лимфоцитов в эффекторные клетки.
С в е т л ы й ц е н т р узелка по строению и функциональному назначению соответствует фолликулам лимфатического узла и является тимуснезависимым участком. Здесь имеются лимфобласты, многие из которых находятся на стадии митоза, дендритные клетки, фиксирующие антиген и сохраняющие его в течение дли тельного времени, а также свободные макрофаги, содержащиепоглощенные продукты распада лимфоцитов в виде окрашенных
телец. |
Строение |
светлого центра |
отражает |
функциональное |
со |
|||
стояние лимфоузелка и может значительно |
изменяться при |
ин |
||||||
фекциях и интоксикациях. Центр окружен |
плотным |
лимфоци- |
||||||
тарным |
ободком — м а н т и й н о й |
з о н о й . |
|
|
|
|
||
Вокруг всего |
узелка располагается |
м а р г и н а л ь н а я |
з о н а , |
|||||
в которой содержатся Т- и В-лимфоциты и макрофаги. |
Полагают, |
|||||||
что в функциональном отношении эта |
зона — один из участков |
|||||||
кооперативного |
взаимодействия |
разных типов клеток |
в |
иммун |
||||
ном ответе. Расположенные в данной зоне В-лимфоциты в резуль тате этого взаимодействия и стимулированные соответствующим антигеном про лифе рируют и дифференцируются в антителообра-
зующие |
плазматические клетки, накапливающиеся в тяжах крас |
|
ной пульпы. |
Форма селезеночного узелка поддерживается с по |
|
мощью |
сети |
ретикулярных волокон — в тимуснезависимом участ |
ке они |
расположены радиально, а в Т-зоне — вдоль длинной оси |
|
центральной артерии.
294 |
295 |
К р а с н а я |
п у л ь п а (pulpa lienis rubra). Обширная часть |
(до 70% массы) |
селезенки, расположенная между лимфатически |
ми узелками и трабекулами. Из-за содержания в ней значитель ного количества эритроцитов имеет па неокрашенных препаратах селезенки красный цвет. Состоит из ретикулярной ткани с нахо
дящимися в ней свободными клеточными элементами: |
клетками |
||
крови, плазматическими клетками и |
макрофагами. |
В красной |
|
пульпе встречаются многочисленные |
артериолы, капилляры и |
||
своеобразные в е н о з н ы е |
с и н у с ы |
(sinus venosus), в их поло |
|
сти депонируются самые |
разнообразные клеточные |
элементы. |
|
Богата синусами красная пульпа на границе с маргинальной зоной лимфатических узелков. Количество венозных синусов в селезенке животных разных видов неодинаково. Их много у кро ликов, морских свинок, собак, меньше у кошек, крупного и мел кого рогатого скота. Участки красной пульпы, расположенные между синусами, называются с е л е з е н о ч н ы м и , или пульпарными тяжами, в составе которых много лимфоцитов и проис ходит развитие зрелых плазматических клеток. Макрофаги пульпарных тяжей осуществляют фагоцитоз поврежденных эритро цитов и участвуют в обмене железа в организме.
Кровообращение. Сложность строения и многофункциональ ность селезенки может быть понята только в связи с особенно стями ее кровообращения.
Артериальная |
кровь направляется |
в |
селезенку |
по |
с е л е з е |
|||||||||
н о ч н о й |
а р т е р и и , |
которая |
через ворота входит |
в орган. От |
||||||||||
артерии |
отходят |
ветви, идущие |
внутри крупных трабекул |
и на |
||||||||||
зывающиеся |
т р а б е к у л я р н ы м и |
а р т е р и я м и . |
В их |
стенке |
||||||||||
имеются |
все |
оболочки, свойственные артериям |
мышечного типа: |
|||||||||||
интима, медия и |
адвентиция. |
Последняя |
срастается с |
соедини |
||||||||||
тельной |
тканью |
трабекулы. От |
трабекулярной |
артерии |
отходят |
|||||||||
артерии |
мелкого калибра, которые вступают в красную |
пульпу и |
||||||||||||
называются |
п у л ь п а р н ы м и |
|
а р т е р и я м и . |
Вокруг |
пульпар- |
|||||||||
иых |
артерий |
образуются удлиненные лимфатические влагалища, |
||||||||||||
по |
мере |
отдаления от |
трабекулы |
они |
увеличиваются и прини |
|||||||||
мают шарообразную форму (лимфатический узелок). Внутри этих лимфатических образований от артерии отходит множество ка пилляров, а сама артерия получает название ц е н т р а л ь н о й . Однако центральное (осевое) расположение имеется лишь в лим фатическом влагалище, а в узелке — эксцентричное. По выходе из узелка эта артерия распадается на ряд веточек — к и с т о ч к о - в ы е а р т е р и о л ы . Вокруг конечных участков кисточковых артериол расположены овальные скопления удлиненных ретику лярных клеток (эллипсоиды, или гильзы). В цитоплазме эндо
телия эллипсоидных |
артериол |
обнаружены |
микрофиламенты, с |
|||
которыми |
связывают |
способность |
эллипсоидов |
сокращаться — |
||
функция |
своеобразных сфинктеров. Артериолы |
далее разветвля |
||||
ются на к а п и л л я р ы , часть |
их |
впадает |
в венозные синусы |
|||
красной пульпы (теория закрытого кровообращения). В соответ ствии с теорией открытого кровообращения артериальная кровь
из капилляров выходит в ретикулярную ткань пульпы, а из нее просачивается через стенку в полость синусов. Венозные синусы занимают значительную часть красной пульпы и могут иметь раз личные диаметр и форму в зависимости от их кровенаполнения. Тонкие стенки венозных синусов выстланы прерывистым эндо телием, расположенным на базальной пластинке. По поверхности стенки синуса в виде колец идут ретикулярные волокна. В конце синуса, на месте перехода его в вену, имеется другой сфинктер.
В зависимости от сокращенного или расслабленного состояния
артериальных |
и венозных сфинктеров синусы |
могут находиться |
в различных |
функциональных состояниях. При |
сокращении ве |
нозных сфинктеров кровь заполняет синусы, растягивает их стен
ку, при этом плазма крови |
выходит через нее |
в ретикулярную |
ткань пульпарных тяжей, а |
в полости синусов |
накапливаются |
форменные элементы крови. |
В венозных синусах селезенки мо |
|
жет задерживаться до 7з общего количества эритроцитов. При открытых обоих сфинктерах содержимое синусов поступает Б; кровоток. Нередко это происходит при резком возрастании по требности в кислороде, когда возникают возбуждение симпатиче ской нервной системы и расслабление сфинктеров. Этому также
способствует сокращепие гладких |
мышц капсулы и трабекул се |
|
лезенки. |
|
|
Отток венозной крови из пульпы происходит по системе вен. |
||
Стенка |
т р а б е к у л я р н ы х в е н |
состоит только из эндотелия,. |
тесно |
прилегающего к соединительной ткани трабекул, то есть |
|
эти вены не имеют собственной мышечной оболочки. Такое строе
ние трабекулярных вен |
облегчает выталкивание |
крови |
из их по |
лости в селезеночную |
вену, выходящую через |
ворота |
селезенки |
и впадающую в воротную вену. |
|
|
|
ГЛАЗА 8 ЭНДОКРИННЫЕ ЖЕЛЕЗЫ
Общая характеристика. Не имеющие выводных протоков и бога то снабженные кровеносными сосудами, эндокринные (endon —
внутрь, krino — отделяю) железы |
вырабатывают биологически! |
активные вещества — г о р м о н ы , |
поступающие непосредственно' |
в кровяное русло, в связи с чем и называют их железами внутрен
ней секреции. Гормоны разносятся кровью по всему |
организму,, |
|
осуществляя гуморальную регуляцию таких важных |
функций,, |
|
как обмен веществ, рост, развитие, |
воспроизводство |
животных;, |
усиливают или замедляют функции |
других систем. |
|
Эндокринные железы в тесной взаимосвязи функционируют* между собой и с нервной системой, образуя при этом единую* нейроэндокринную систему. Эта система призвана поддерживатьпостоянство (гомеостаз) организма и обеспечивать его связь с, внешней средой.
296 |
2977 |
Гормоны одной железы по принципу обратной связи воздей
ствуют на функцию |
другой железы, оказывая влияние только |
на клетки — мишени. |
Последние характеризуются тем, что в их |
плазмолемме располагаются специфические химические рецеп торы.
По химическому составу гормоны являются аминокислотны ми группами, гликопротеидами, стероидами (производные холе стерина).
Эндокринные железы построены из эпителиальной или нерв ной ткани. Эпителиальные клетки образуют пузырьки (фолли кулы) или тяжи. Такая ориентация создает наилучшие условия контакта гормонопродуцирующих клеток и кровеносных сосудов. Нейроны гипоталамуса в ответ на воспринятое раздражение с помощью гранулярной эндоплазматической сети вырабатывают биологически активные вещества — н е й р о г о р м о и ы (нейросекреты). Они оттекают по аксону и, поступая в кровь через аксовазальные (сосудистые) синапсы, усиливают или замедляют гу моральным путем функцию клеток. Нейрогормон — специфиче ский эффекторный сигнал нейросекреторной клетки в ответ на афферентную импульсацию.
Таким образом, нейросекреториые нейроны — это промежу точное звено между эндокринными железами и центральной нервной системой.
Железы внутренней секреции развиваются из эктодермы и эмбриональных зачатков нервной системы.
Классификация. К железам внутренней секреции относят нейросекреторные ядра гипоталамуса, гипофиз, эпифиз, щитовид ную железу, околощитовидную железу, надпочечники; эндокрин ную часть поджелудочной железы, яичника, семенника, почки, плаценты; одиночные гормоносинтезирутощие клетки неэндо кринных органов и тканей. Эти железы формируют нейроэндокрипную систему, в которой в настоящее время выделяют два взаимосвязанных звена: центральное и периферическое.
Центральное звено — это нейросекреторные ядра гипоталаму са, гипофиз, эпифиз. В периферическое звено входят железы, функция которых зависит от передней доли гипофиза (щитовид ная железа, кора надпочечников, семенники, яичники) и желе8ы, функция которых не зависит от передней доли гипофиза (мозговое вещество надпочечников, околощйтовидная железа, околофолликулярные — кальцитонипоциты щитовидной железы, гормоносинтезирующие клетки неэндокринных органов.
, ЦЕНТРАЛЬНЫЕ ЗВЕНЬЯ ЭНДОКРИННОЙ СИСТЕМЫ
ГИПОТАЛАМУС
Гипоталамус — подбугровая область промежуточного мозга. Обра зует стенку и дно третьего мозгового желудочка (рис. 214). Нерв ные клетки серого вещества гипоталамуса формируют 32 пары
298
ядер, распределенные в трех зонах: передней, средней, задней. Нейросекреторные клетки этих ядер вырабатывают биологически активные вещества — нейрогормоиы, которые в отличие от медиа торов характеризуются дистантным и более ' продолжительным действием.
В переднем гипоталамусе расположены парные супраоптические и паравентрикулярные ядра (рис. 215). Они состоят иа крупных нейросекреторных клеток, цитоплазма и отростки кото рых содержат секреторные гранулы. Эти клетки имеют хорошо развитую гранулярную эндоплазматическую сеть; с ее участием протекает образование белковых нейрогормонов: вазопрессина и окситоцина. Оттекая по аксонам и накапливаясь в виде нако пительных телец в задней доле гипофиза (нейрогипофизе), они поступают в дальнейшем в кровь.
Вазопрессин суживает просвет кровеносных сосудов и повыг шает кровяное давление; регулирует водный обмен, так как влияет на обратное всасывание воды в мочевых канальцах почки.
Окситоцин стимулирует функцию гладких мышц матки, спо собствуя этим выделению секрета мочеточных желез; природах вызывает сильное сокращение мышечной оболочки стенки матки; влияет на сокращение мышечных элементов молочной железы.
Крупные нейросекреторные клетки супраоптических и паравентрикулярных ядер ввиду их принадлежности к парасимпати ческому гипоталамусу относят к холинергическим нейронам.
Между ядрами переднего гипоталамуса и нейрогипофизом существует тесная морфофункциональная связь, что позволяет выделять гипоталамонейрогипофизарную систему в едином гипоталамогипофизарном комплексе.
299

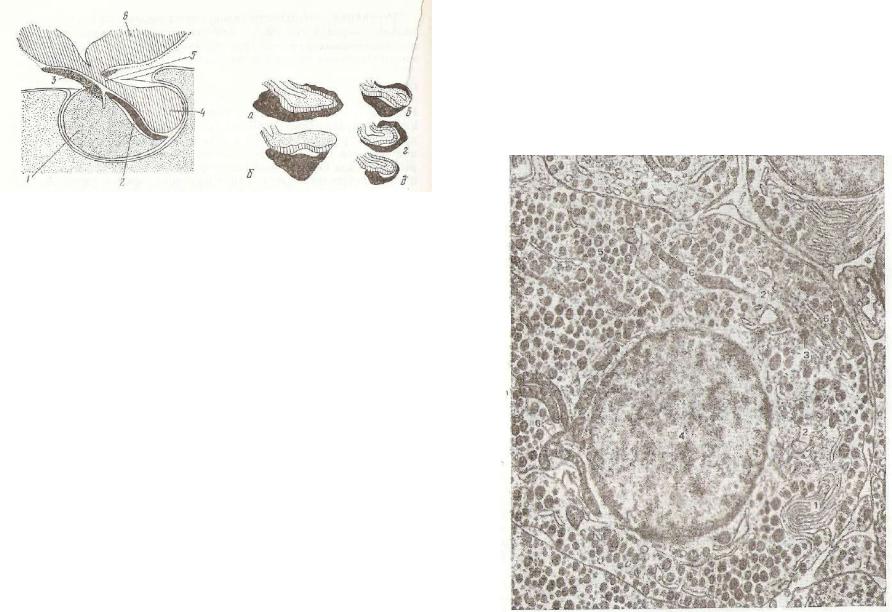
Рис. 216. Строение и изменение в процессе секреции структур ных компонентов перикариоп^ нейросекреторной клетки ния/- небугорной области (гипотала муса), ее отростков и терми нален, контактирующих с эпи телиальной железистой клет кой (18) и с капилляром (29):
1 — |
свободные рибосомы; |
2 — |
|||
гранулярная |
эндоплазматическая |
||||
сеть, |
содержащая первичные се |
||||
креторные продукты; 3 — грану |
|||||
лярная |
секреторная сеть бее пер |
||||
вичного |
секреторного |
продукта; |
|||
4 — |
переходный элемент |
эндо- |
|||
плазматической сети; 5 — неак |
|||||
тивные мембранные и вакуолярные |
|||||
компоненты |
аппарата |
Гольджи; |
|||
6 — те же компоненты, конденси |
|||
рующие |
секреторный |
продукт; |
|
7 — зрелые гранулы нейросекре- |
|||
та; 8 — лизосомоподобные круп |
|||
ные гранулы; |
9 — митохондрии; |
||
10 — нейрофиламенты; 11 — оста |
|||
точные нейросекреторные |
грану |
||
лы; 12 — синаптические пузырь |
|||
ки; 13 — |
пре- и постсинаптиче- |
||
ская мембрана; 14 — мелкие гра |
|||
нулы, содержащие катехоламины; |
|||
15 — базальная мембрана капил |
|||
ляров; 16 — микроворсинки; 17— |
|||
эритроцит; |
20 |
— синаптические |
|
бляшки. |
|
|
|
В ядрах медиобазального гипоталамуса расположены мелкие нейросекреторные клетки, продуцирующие группу тропных нейрогормонов. Они накапливаются в медиальной эминенции, где затем поступают в кровь и в дальнейшем по портальной сосуди стой системе транспортируются в аденогипофиз, усиливая или ослабляя его гормонообразование. Действие этих гормонов на правлено на регуляцию функций щитовидной железы, коры над почечников, половых органов.
Всвязи с принадлежностью мелких нейросекреторных клеток
ксимпатической части гипоталамуса их относят к адренергическим нейронам. Наряду с нейросекреторными клетками здесь находятся и обычные нейроны адренергического типа, которые синтезируют и выделяют медиаторы, действующие локально в зоне их синапсов. Эти нейроны осуществляют связь между дру гими ядрами внутри гипоталамуса, а также дают начало нисхо дящим нервным волокнам, идущим в стволовую часть головного мозга и в спинной мозг.
Существующая тесная морфофункциональная связь медиобазальной части гипоталамуса и аденогипофиза обусловила выде ление еще одной гипоталамоаденогипофизарной системы единого гипоталамогипофизарного комплекса (рис. 216).
Нейросекреторная функция гипоталамуса регулируется норадреналином, серотонином, ацетилхолином. Эти нейроамины син тезируются в зонах центральной нервной системы, не связанных с гипоталамусом. Функции гипоталамуса регулируются также симпатической нервной системой и гормонами эпифиза.
Регуляция деятельности эндокринных желез и их связей с цент ральной нервной системой —это лишь одна из многих важных интеграционных функций гипоталамуса, который является в орга низме животных высшим вегетативным центром.
ГИПОФИЗ
Гипофиз — компонент единой гипоталамофизарной системы орга низма. Вырабатывает гормоны, регулирующие функцию многих желез внутренней секреции и осуществляет их связь с централь ной нервной системой. Расположен он в гипофиз арной ямке ту рецкого седла клиновидной кости черепа; имеет бобовидную форму и очень небольшую массу. Так, у крупного рогатого скота она око ло 4 г, а у свиней меньше — 0,4 г.
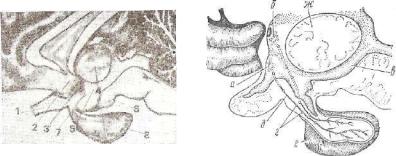
Развивается гипофиз из двух эмбриональных зачатков, расту щих навстречу друг другу. Первый зачаток — гипофизарный кар ман — образуется из крыши первичной ротовой полости и направ лен в сторону головного мозга. Это эпителиальный зачаток, из которого в дальнейшем развивается аденогипофиз.
Второй зачаток — выпячивание дна мозгового желудочка, по этому он является мозговым карманом и из него образуется нейрогипофиз (рис. 217).
Эмбриогенез обусловил строение органа — гипофиз состоит из двух долей: аденогипофиза и нейрогипофиза (рис. 218, 219).
В состав аденогипофиза входят передняя, промежуточная и туберальная части. Передняя часть построена из эпителиальных клеток — аденоцитов, формирующих тяжи (трабекулы) и разгра ниченных синусоидными капиллярами вторичной сосудистой сети (цв. табл. VII—А—а). Первичная сосудистая сеть расположена в медиальной эминенции.
А — ранняя и В — более поздняя стадии; а — стенки нервной трубки; б — стенки мозгового пузыря; в — эпителий ротовой бухты; г — хорда; д — кишечная трубка; е — мезенхима; ж — гипофизарный карман; его а — передняя и и — задняя стен ки; к — зачаток задней части.
300 |
301 |
|
Соединительнотканная строма аденогипофиза развита слабо. Аденоциты по-разному воспринимают красители: клетки, хоро
шо окрашивающиеся, называют хромофильными, а плохо крася щиеся— хромофобными (б). Хромофильные аденоциты могут вос принимать или кислые красители или основные, поэтому первые именуют ацидофильными (в), вторые — базофильными (г).
Ацидофильные клетки составляют 30—35% всех клеток перед ней части гипофиза. Они имеют округлую или овальную форму, крупнее хромофобных и мельче базофильных аденоцитов. В цито плазме ацидофила содержатся гранулы, окрашивающиеся эозином; ядро расположено в центре клетки. К нему прилегают комплекс Гольджи, незначительное количество крупных митохондрий, хо рошо развита зернистая эндоплазматическая сеть, что свидетель ствует об интенсивном синтезе белка.
В связи с разной гормонообразовательной функцией и строе нием, цитоплазматической зернистостью различают три разновид ности ацидофильных аденоцитов: соматотропоциты, лактотропоциты, кортикотропоциты. Соматотропоциты продуцируют соматотропный гормон, стимулирующий рост тканей и всего организма в це лом. Лактотропоциты образуют пролактин (лактотропный гормон), регулирующий процесс лактации и функциональное состояние желтого тела яичника. Кортикотропоциты вырабатывают кортикотропин, повышающий гормонообразовательную функцию коры зва дпочечников.
Секреторные гранулы |
соматотропоцитов шаровидной |
формы, |
|
в диаметре от 200 до 400 |
нм (рис. |
220). Лактотропоциты имеют |
|
€олее крупные секреторные гранулы |
овальной формы с |
длиной |
|
500—600 нм, шириной 100—120 нм. Секреторные гранулы кортикотропоцитов снаружи покрыты мембраной пузырчатой формы е
плотной сердцевиной. |
- ЩЩ |
302 |
|
Базофильные аденоциты составляют 4—10% всех клеток пе редней части гипофиза. Это самые крупные клетки аденогипофи за. Их секреторные гранулы имеют гликопротеидный характер, поэтому окрашиваются основными красителями. Различают две разновидности этих клеток: гонадотропные и тиротропные. Гонадотропные клетки продуцируют фолликулостимулирующий гор мон, регулирующий развитие женских и мужских половых клеток, секрецию половых органов самки и лютеинизирующего гормона, стимулирующего рост и развитие желтого тела в яичниках и ин-
303