

gistologia
.pdfРис. 155. Вазофильная глыбчатостьв |
Рис. 156. Электронная микрофото- |
||||||
нейроплазме перикарионов узловато- |
грамма глыбок тигроидного вещества |
||||||
го ганглия блуждающего нерва кро- |
нервной клетки (ув. 84200). |
|
|||||
лика (препарат Н. А. Козлова): |
|
|
|
|
|||
1 — глыбки Ниооеля; |
2 — аксонный |
|
|
|
|||
холмик (место отхождения аксона); 3— |
|
|
|
||||
ядра глиоцитов. |
|
|
|
|
|
|
|
ных |
цистерн |
гранулярной |
эндоплазматической |
сети |
|||
(рис. |
156). |
|
|
|
|
|
|
Высокое содержание |
гранулярной эндоплазматической сети в |
||||||
нейронах соответствует |
их |
высокому уровню |
синтеза белков в |
||||
связи |
с необходимостью |
поддерживать массу |
цитоплазмы пери |
||||
карионов нервных |
клеток и их длинных отростков. Масса веще |
||||||
ства аксона, не имеющего |
рибосом, поддерживается постоянным |
||||||
током |
цитоплазмы от тела |
клетки в периферический отдел от |
|||||
ростка со скоростью от 1 мм и выше в сутки, что обеспечивает их целостность и функциональную активность.
Степень плотности и ориентации цистерн гранулярной эндо плазматической сети специфична для различных отделов нерв ной системы. Например, в моторных нейронах спинного мозга глыбки базофильного вещества крупные и угловатые. Они плот нее локализованы вокруг ядра, мельче и лежат реже на перифе рии клетки. В чувствительных клетках спинальных ганглиев глыбки мелкие в виде пылевидной зернистости и т. д.
В эмбриональном периоде развития объем цитоплазмы ней ронов резко увеличивается, возрастает в их цитоплазме содержа ние РНК и соответственно увеличивается масса базофильного вещества.
204
К о м п л е к с |
Г о л ь д ж и при световой микроскопии виден |
как совокупность |
извивающихся нитей, колечек и зернышек в |
средней зоне цитоплазмы перикариона клетки. Под электронным |
|
микроскопом выявляют типичные компоненты этой органеллы: группы плоских цистерн, вакуоли и микропузырьки.
М и т о х о н д р и и мелкие, палочковидной формы. Расположе ны как в перикарионе, так и отростках, в том числе и в межнепрональных синапсах.
Ц е н т р о с о м а состоит из двух центриолей. Наличие клеточ ного центра в настоящее время установлено в нейронах почти всех отделов нервной системы.
НЕЙРОГЛИЯ
Нейроглия — комплекс клеточных элементов, выполняющих в нервной ткани опорную, разграничительную, трофическую, сек реторную и защитную функции. В составе нейроглии различают макроглшо п микроглию.
Глиоцпты макроглии развиваются в нервной трубке одновре менно с нейронами. В составе глиоцитов различают эпендимоциты, астроциты и олигодеидроциты. Э п е н д и м о ц и т ы выстилают полости центральной нервной системы: желудочки головного моз га п спинно-мозговой канал. Кубические или призматические клетки эпендимы на поверхности, обращенной в полость нервной трубки, содержат реснички. Их противоположные полюса пере ходят в длинные отростки. Последние включаются в поддержи вающий остов тканей нервной трубки. Достигая ее внешней по верхности или стенок кровеносных сосудов, они включаются в со-
а — эпендимные клетки с мерцательными и базальнымп отростками (Ь), с — рлиальная пограничная мембрана, образованная астроцитами (d), с отростками кото рых переплетаются отростки эпендимных клеток.
205
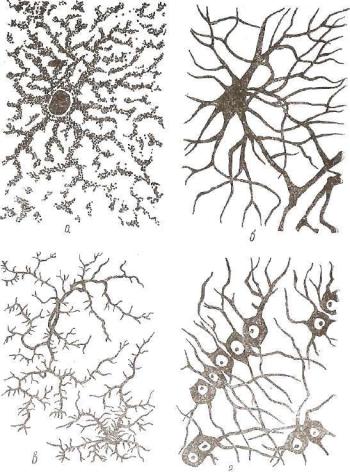
а — плазматические астроциты; б — волокнистые астроциты- |
|
дендроглиоциты; г — глиальные макрофаги. |
' |
став разграничительных мембран, формирующихся между нервной и другими тканями центральной нервной системы. Эпендимоциты участвуют и в секреторной функции, выделяя в полости централь ной нервной системы или в кровь различные активные вещества (рис. 157).
А с т р о ц и т ы бывают коротколучистые (протоплазматические) и длиннолучистые (волокнистые). Первые локализованы в сером веществе центральной нервной системы. Морфологически они характеризуются большим, бедным хроматином ядром и мно гими короткими, сильно ветвящимися отростками (рис. 158). Их
206
цитоплазма бедна цистернами эндоплазматической сети. Она со держит мало свободных рибосом, но богата митохондриями. Плаз матические астроциты участвуют в обмене веществ нервной ткани и выполняют разграничительную функцию.
Волокнистые астроциты характерны для белого вещества центральной нервной системы. Они имеют небольшой по объему перикарион и 30—40 тонких, длинных, переходящих в глиальные волокна отростков. Цитоплазма клеток прозрачна и содержит многочисленные фибриллы 8—9 нм в диаметре.
Волокна астроглии на кровеносных сосудах и поверхности мозга формируют разграничительные мембраны. Волокнистые астроциты образуют поддерживающий аппарат центральной нерв ной системы.
О л и г о д е н д р о г л и о ц и т ы — большая |
разнообразная |
по |
форме-и специфичности функционального |
значения группа |
кле |
ток центральной и периферической нервной системы. Они окру жают тела нейронов, входят в состав оболочек нервных волокон и нервных окончаний (леммоциты, или шванновские клетки),
участвуют в их обмене |
веществ. |
||
Микроглия |
(глиальные |
макрофаги) — специализированная |
|
•система макрофагов, |
тканей |
внутренней среды, выполняющая |
|
защитную функцию. Развиваются они из мезенхимы. Форма кле ток отростчатая и в соответствии со способностью клеток и аме боидному перемещению непостоянна. Их ядра богаты хромати ном, вытянутой или неправильной формы. При активизации клет ки округляются. Они характерны для белого и серого вещества центральной нервной системы.
НЕРВНЫЕ ВОЛОКНА
Отростки нервных клеток в совокупности с покрывающими их клетками нейроглии образуют нервные волокна. Расположенные в них отростки нервных клеток (дендриты или нейриты) назы вают осевыми цилиндрами, а покрывающие их клетки олигодендроглии — и е й р о л е м м о ц и т а м и (леммоцитами, шванновскими клетками). В соответствии с составом нервных волокон и морфологическими особенностями их строения различают миелииовые и безмиелииовые нервные волокна.
Безмиелиповые (безмякотные) нервные волокна характерны для вегетативной нервной системы. Леммоциты — клетки олигодепдроглтти — в составе безмиелипового волокна плотно приле тают друг к другу, образуя непрерывные тяжи. При световой микроскопии грапицы глиальных клеток в волокне не просмат риваются и клетки в совокупности выглядят как непрерывная лента, содержащая характерные для них овальные ядра. В безмякотном нервном волокне содержится несколько осевых ци линдров, то есть отростков различных нервных клеток, которые -свободно могут покидать его ж переходить в смежные волокна. При формировании нервного волокна плазмолемма глиальной.
207

Рис. 160. Схема строения безмиелинового нервного волокна: |
|
1 — ядро и 2 — цитоплазма леммоцита; 8—4 — аксоны нервных клеток, погружен |
|
ные в цитоплазму леммоцита (8 — полностью, 4 — частично); |
5 — митохондрия; |
6 — шероховатая эндоплазматическая сеть; 7 — микротрубочки; |
в — коллагеновые |
волокна эндоневрия; 9 — базальная мембрана; 10 — мезаксон. |
|
I I L
1 — ядро леммоцита; 2 — цитоплазма леммоцита; 3 — осевые цилиндры; 4 — ме* заксон (по Фаусетту).
клетки прогибается, образуя более или менее глубокие складки,, содержащие отростки нервных клеток — осевые цилиндры. Сбли женные в области складки участки плазмолеммы леммоцита формируют сдвоенную мембрану — м е з а к с о н , на котором под вешен осевой цилиндр. При световой микроскопии плазмолемма леммоцита и мезаксоны не выявляются, что создает впечат ление погружения осевых цилиндров непосредственно в цито плазму глиальных клеток (рис. 159, 160, 161).
Миелиновые (мякотные) нервные волокна. Диаметр миелиновых волокон колеблется от 1 до 20 мкм. Они содержат один осе вой цилиндр — дендрит или нейрит нервной клетки, покрытый оболочкой, образованной последовательно расположенными клет ками иейроглии — леммоцшами.. В оболочке волокна различают
]4 Заказ № 908 |
20& |

Рис. 162. Миелиновые нервные волокна из седалищ ного нерва лягушки, обработанные осмиевой кисло той:
1 — осевой цилиндр; 2 — леммоцит; 3 — миелиновая обо^ лочка; 4 — соединительная ткань; б — насечки; б — перехват.
два слоя: внутренний — миелиновый, более толстый и наружный — тонкий, содержа щий цитоплазму и ядра леммоцитов.
На границе двух леммоцитов оболочка миелинового волокна истончается, образуется сужение волокна — у з л о в о й п е р е х в а т (перехват Ранвье). Участок нервного волок на между двумя узловыми перехватами на зывается межузловым сегментом. Его оболоч ка соответствует одному леммоциту (рис. 162).
В процессе развития миелинового нервно го волокна осевой цилиндр, погружаясь в ци топлазму леммоцита, вовлекает с собой его плазмолемму. Здесь, как и в безмиелиновом волокне, формируется мезаксон, который, по~ следовательно удлиняясь и концентрически наслаиваясь на осевой цилиндр, образует зо ну плотно расположенных мембран — миелиновую оболочку волокна (рис. 163, 164).
При электронной микроскопии видно, что
.мембраны мезаксона образуют чередующиеся светлые линии (8— 12 нм), соответствующие их липидным слоям, и темные тонкие, образованные белковыми молекулами (рис. 165).
Наружный слой оболочки миелинового волокна — н е в р и- л е м м а (шванновская оболочка) — соответствует оттесненной на периферию цитоплазме леммоцитов с их ядрами. Плотно распо-
.ложенные витки мезаксона в соответствующих участках волок- •на, отклоняясь от параллельной осевому цилиндру ориентации, рассредоточиваются и формируют косо ориентированные зоны -оболочки, более богатые цитоплазмой. При осмировании волокна они выделяются как светлые линии — насечки миелина.
В области перехвата слои мембран миелиновой оболочки контактируют с осевым цилиндром волокна. В соответствии с последовательностью роста осевого цилиндра и формирования •слоев мезаксона более глубокие слои последнего короче поверх ностных и располагаются дальше от перехвата.
Смежные леммоциты оболочки волокна взаимодействуют друг •о другом системой более или менее выраженных пальцевидных отростков, которые, переплетаясь, формируют их контакты и на •срезах наблюдаются в различных сечениях. Миелиновое волокно с поверхности покрыто базальной мембраной, связанной с тяжа ми коллагеновых волокон окружающей соединительной ткани.
510
Рис. 163. Схема развития миелинового волокна:
1 — контакт аксолеммы и оболочки леммоцита; 2 — щель; з — аксолемма и обо лочки леммоцита; 4 — цитоплазма леммоцита; 5 — мезаксон.
Рпс. 164. Схема строения миелинового волокна:
1 — аксон; 2 — мезаксон; з — насечки; 4 — перехват; S — протоплазма леммоцита; в — ядро леммоцита; 7 — неврилемма; 8 — эндоневрий.
Рис. 165. Электронная ар|{рофотография поперечного среза миелинового во локна из краниального шейного узла крупного рогатого скота (препарат Козлова):
1 — ядро леммоцита; 2 — цитоплазма.леммоцита; з — пластины миелиновой обо лочки; 4 — нейрофиламентыв цитоплазме аксона.
14*
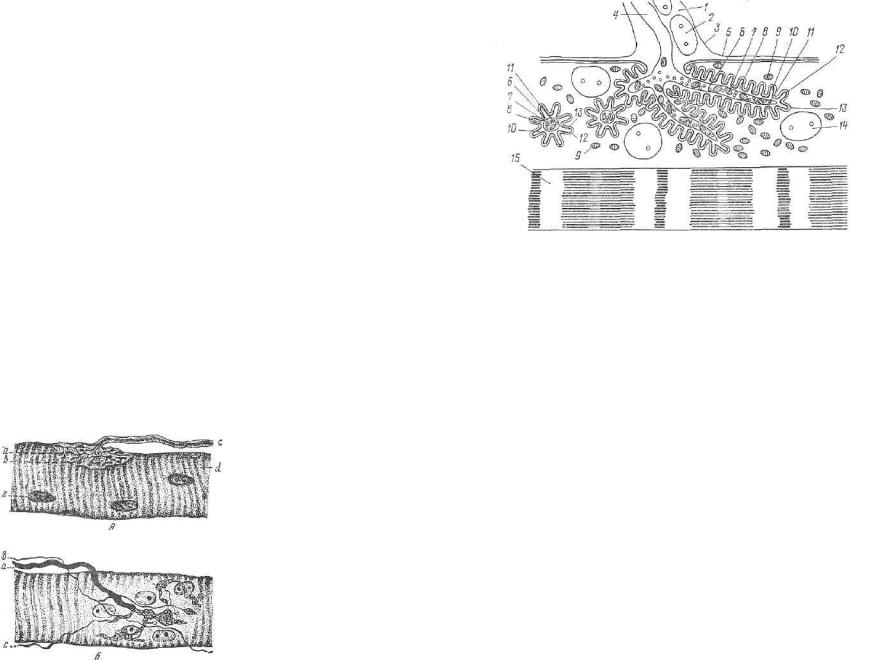
Осевой цилиндр нервных волокон состоит из нейроплазмы — ци топлазмы нервных клеток, содержащей продольно ориентирован ные нейрофиламенты и нейротубулы. Наличие различных оргапелл и их локализация специфичны для осевых цилиндров во локон различного функционального значения.
НЕРВНЫЕ ОКОНЧАНИЯ
Концевые аппараты нервных волокон — нервные окончания — различны по их функциональному значению. Различают три вида нервных окончаний: эффекторные, рецепторные и концевые ап параты в составе межнейрональных синапсов.
Эффекторные нервные окончания — к их числу относят дви гательные нервные окончания поперечнополосатых и гладких мышц и секреторные окончания железистых органов.
Д в и г а т е л ь н ы е н е р в н ы е о к о н ч а н и я п о п е р е ч н о
п о л о с а т ы х с к е л е т н ы х м ы ш ц — м о т о р н ы е |
б л я ш |
ки — комплекс взаимосвязанных структур нервной и |
мышечной |
тканей. Моторная бляшка — эффекторный аппарат аксонов нерв ных клеток двигательных ядер передних рогов спинного мозга или двигательных ядер головного мозга и мышечных волокон. Морфологически он состоит из нервного полюса — терминальной части аксона нейрона и мышечного полюса, специализированного
участка мышечного волокна — подошвы моторной |
бляшки |
(рис. 166). |
|
Моторное нервное волокно вблизи мышечного волокна утра чивает сопровождающие осевой цилиндр ядра глиальных клеток и миелиновую оболочку. Осевой цилиндр, распадаясь на несколь ко терминальных веточек, погружается в специализированную вишу мышечного волокна.
Сакролемма в области нервного окончания образует много численные субмикроскопические складки, формирующие вторич ные синаптические щели мо
торного окончания.
Мышечное волокно в об ласти подошвы моторной бляшки не имеет миофибрилл и поперечной исчерченно-
Рис. 166. Двигательное нервное
окончание (моторная бляшка}:
А — вид в профиль (а и Ъ — окон чания миелинового нервного волок на, с — миелиновое волокно, d — мышечное волокно, е — ядро мышеч ного волокна); В — вид сверху (а — миелиновое волокно, Ъ — безмиелиновое нервное волокно, с — волок но, выходящее из моторной бляшки и оканчивающееся в другой мотор ной бляшке, так называемое «уль тратерминальное волокно»).
Рис. 167. Схема строения моторной бляшки:
I — питоплазма леммоцита; 2 — ядро; з — неврилемма; 4 — осевой цилиндр; 5 — сарколемма; 6 — концевые веточки нервного волокна в продольном и поперечном сечениях; 7 — митохондрии в нейроплазме (аксоплазме); 8 — первичное синаптическое пространство; 9 — саркосомы; ю — вторичное синаптическое пространство; II — синаптические пузырьки; 12 — пресинаптическая мембрана; 13 — постсинаптическая мембрана; 14 — ядро моторной бляшки (мышечное); 15 — миофибрилла, состоящая из миопротофибрилл.
сти. Здесь цитоплазма содержит значительное количество мито хондрий и круглых или овальных ядер. Совокупность этих струк тур мышечного волокна в области нервного окончания образует его мышечный полюс.
Терминальные ветви |
осевого цилиндра нервного волокна ха |
|
рактеризуются наличием |
митохондрий и многочисленных синап- |
|
тических пузырьков, |
содержащих |
медиатор — ацетилхолин |
(рис. 167). Последний при деполяризации плазмолеммы аксо на — пресинаптической мембраны — поступает в синаптическую щель и на холинорецепторы постсинаптической мембраны, ко торой служит оболочка мышечного волокна, что вызывает воз буждение (волну деполяризации постсинаптической мембраны).
М о т о р н ы е н е р в н ы е о к о н ч а н и я г л а д к о й мы
ш е ч н о й т к а н и |
формируются нервными волокнами, которые |
|
распространяются |
между мышечными клетками и образуют чет- |
|
кообразные |
расширения, содержащие холинергические или адре- |
|
нергические |
пузырьки. |
|
Чувствительные нервные окончания (рецепторы) — специали зированные концевые образования дендритов чувствительных нейронов. В соответствии с их локализацией и специфичностью участия в нервной регуляции жизнедеятельности организма раз личают две большие группы рецепторов: экстерорецепторы и интерорецепторы. В зависимости от характера воспринимаемого раздражения чувствительные окончания делятся на механорецепторы, хеморецепторы, терморецепторы и др.
212 |
213 |
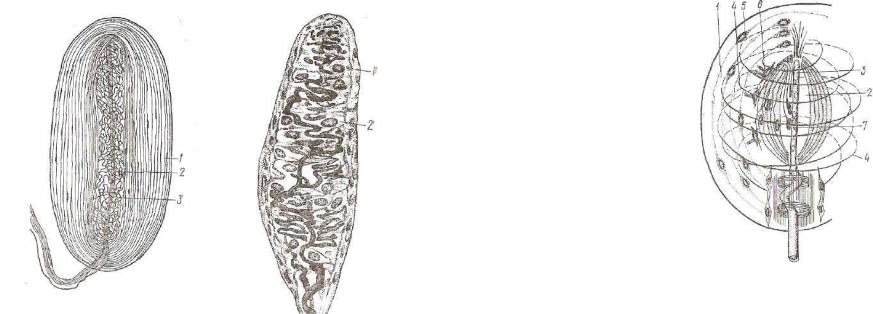
Рис. 168. Пластинчатое тельце (тель- |
|
Рис. 169. Осязательное (ней- |
||||||||
це Фатера—Пачини): |
|
|
|
|
снерово) тельце: |
|
|
|||
1 — наружная |
колба; |
2 — внутренняя |
|
1 — капсула; 2 — специаль- |
||||||
колба; з — концевой отдел нервного во- |
|
ные клетки, |
|
|
|
|||||
локна (по Клара). |
|
|
|
|
|
|
|
|
||
Чувствительные нервные окончания весьма разнообразны по» |
||||||||||
их структурной |
организации. |
Они |
подразделяются на |
с в о б о д |
||||||
н ы е |
нервные |
окончания, состоящие |
только из |
терминальных |
||||||
ветвей |
дендрита чувствительной клетки, и н е с в о б о д н ы е , |
со |
||||||||
держащие в своем составе клетки глии. |
Несвободные окончания,, |
|||||||||
покрытые соединительнотканной |
капсулой, называются |
и н к а п |
||||||||
с у л и р о в а н н ы м и . |
Примером |
свободных нервных окончаний |
||||||||
могут |
служить |
терминальные |
ветвления дендритов |
чувствитель |
||||||
ных клеток в эпидермисе кожи, |
где |
чувствительные |
нервные |
|||||||
волокна, проникая в эпителиальную |
ткань, распадаются на |
тон |
||||||||
кие терминальные веточки. |
|
|
|
|
|
|
|
|||
Весьма разнообразны чувствительные окончания в соедини тельной ткани животных, которые представлены двумя группа ми: н е и н к а п с у л и р о в а н н ы х и и н к а п с у л и р о в а н н ы х нервных аппаратов. Первые содержат ветвящийся осевой цилиндр волокна, сопровождающий глии. Вторые характеризуют ся наличием соединительнотканной капсулы и специфичностью морфологии и функций их глиальных элементов. К группе таких чувствительных окончаний относят пластинчатые тельца (тельца Фатера — Пачини), осязательные тельца (тельца Мейснера), генитальные тельца и др. (рис. 168, 169).'
214
«Рис. 1170. Схема строения пластинчатого тельца:
1 —'слоистая капсула; 2 — внутренняя колФа; з — дендрит чувствительной нервной клетки; 4 — спиральные коллагеновые во локна; 5 — фиброциты; б — глиальные клет ки с ресничками; 7 — синаптические кон такты аксонов вторично чувствующих кле ток с дендритами чувствительной нервной клетки (по Отелину).
В составе |
п л а с т и н ч а т о г о |
|
||
т е л ь ц а различают |
внутреннюю |
|
||
колбу и капсулу. Внутренняя кол- |
|
|||
•ба образована специализированны |
|
|||
ми леммоцитами. В нее погружен |
|
|||
осевой |
цилиндр — терминальный |
|
||
отдел |
чувствительного |
нервного |
|
|
волокна. Проникая во внутреннюю |
|
|||
колбу, он распадается на тончай |
|
|||
шие конечные ветвления. |
|
|||
Капсула |
пластинчатого тельца состоит из |
большого числа |
||
соединительнотканных |
пластинок, образованных |
фибробластами |
||
л спирально |
ориентированными пучками коллагеновых волокон. |
|||
На границе |
наружной капсулы и внутренней колбы лежат клет |
|||
ки, которые |
предположительно определяют как глиальные. Они |
|||
образуют синапсы с ветвлениями осевого цилиндра (рис. 170). Предполагают, что нервный импульс генерируется в условиях •смещения наружной капсулы по отношению к внутренней кол
бе. Пластинчатые тельца характерны для |
глубоких слоев кожи и |
внутренних органов. |
|
О с я з а т е л ь н ы е т е л ь ц а также |
образованы клетками |
тлии, которые ориентированы перпендикулярно к длинной оси
тельца, и стелются по их поверхности терминальными |
ветвями |
|||
•аксона. С поверхности тельце покрыто тонкой |
соединительно |
|||
тканной капсулой. |
|
|
|
|
Аналогично построены и |
г е н и а л ь н ы е т е л ь ц а |
половых |
||
•органов. Отличительной особенностью |
этого вида |
окончаний яв |
||
ляется то, что в генитальное |
тельце |
под капсулу |
проникает не |
|
один осевой цилиндр, а несколько. Последние ветвятся между глиальными клетками тельца. По той же схеме построены кол
бы |
Краузе, с функцией которых связывают температурную чув |
||||
ствительность. При |
их возбуждении |
медиатор поступает в синап- |
|||
тическую щель |
на |
холинорецепторы |
постсинаптической мембра |
||
ны |
мышечного |
волокна и вызывает |
импульс (волну |
деполяри |
|
зации) . |
|
|
|
|
|
|
Р е ц е п т о р ы |
с к е л е т н ы х м ы ш ц — мышечные |
веретена |
||
содержат несколько |
интрафузальных мышечных волокон, покры |
||||
тых общей соединительнотканной капсулой. В состав веретена обычно входят два толстых центральных мышечных волокна и
215
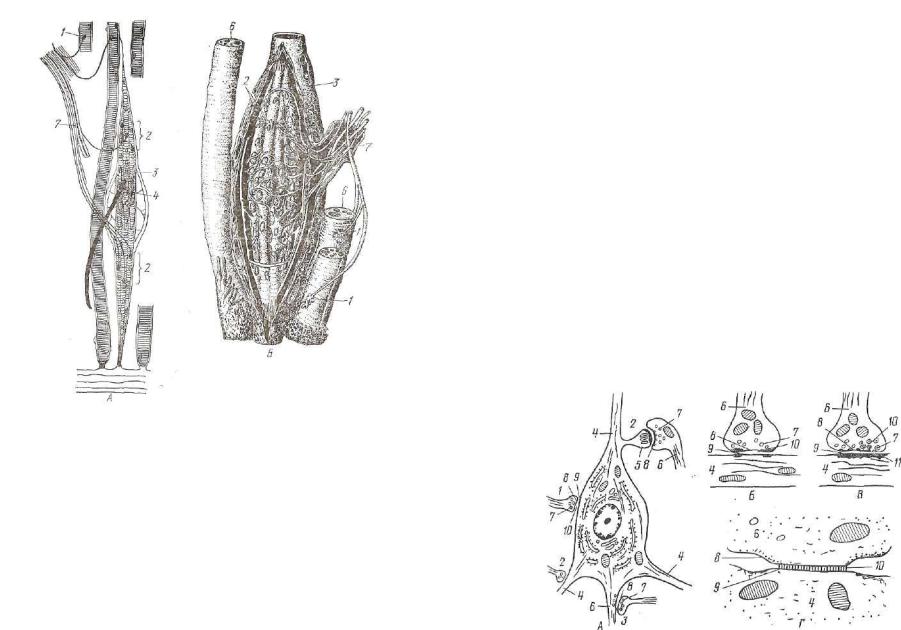
А
Рис. 171. Схема строения нервно-мышечного веретена:
А —• моторная иннервация интрафузальных и экстрафузальных мышечных волоков (по Студитскому); Б — спиральные афферентные нервные окончания вокруг интра фузальных мышечных волокон в области ядерных сумок (по Кристичу с измене нием); 1 — моторные бляшки экстрафузальных мышечных волокон; 2 — моторные бляшки интрафузальных мышечных волокон; 3 — ядерная сумка; 4 — ядерная сумка; 5 — чувствительные аннулоспиральные нервные окончания вокруг ядерныу сумок; 6 — поперечнополосатые мышечные волокна; 7 — нерв.
четыре # тонкжх (рис. 171). Экваториальная часть толстых волокон заполнена скоплениями ядер — «ядерная сумка». В тонких мы шечных волокнах ядра располагаются цепочкой, образуется ядер ная цепь. Чувствительные нервные волокна представлены здесь двумя видами. Одни образуют спиральные завитки, окружающие экваториальную, содержащую ядра, часть толстых интрафузаль ных мышечных волокон—«аннулярные окончания». Окончания второй группы чувствительных волокон представлены как аннулярными окончаниями, так и вторичными гроздьевидными окон чаниями по одному с каждой стороны от первичного. Окончания первой группы реагируют на степень растяжения мышц и на его скорость, вторичные — только на степень растяжения. На обоих полюсах мышечных волокон локализуются окончания дви-
216
гательных нервных волокон и имеют строение, типичное для мо торной бляшки.
Межнейрональный синапс — специализированный контакт двух нейронов, обеспечивающий одностороннее проведение нерв ного возбуждения. Морфологически в синапсе различают пресинаптический полюс — концевой отдел первого нейрона, и пост-
синаптический |
полюс — область контакта |
второго нейрона с пре- |
||
синаптическим |
полюсом первого. Встречаются синапсы с хими |
|||
ческой |
и электрической передачей. |
|
|
|
По месту контакта преганглионарного |
нервного |
волокна с |
||
вторым |
нейроном бывают а к с о с о м а т и ч е с к и е |
с и н а п с ы |
||
(аксон первого нейрона контактирует с перикарионом второго),
а к с о д е н д р и т и ч е с к и е |
(аксон первого нейрона взаимодейст |
|
вует с дендритом второго) |
и а к с о а к с о н а л ь н ы е |
(аксон од |
ного нейрона оканчивается на аксоне другого) (рис. |
172). Пред |
|
полагается, что последний |
не возбуждает нервного импульса на |
|
втором нейроне, а тормозит возбуждение, полученное нейроном через другие синапсы.
Морфологически п р е с и н а п т и ч е с к и й п о л ю с синапса характеризуется наличием синаптических пузырьков, содержа щих медиатор (ацетилхолин или норадреналин), митохондрий, одиночных цистерн, иногда нейротубул. Участие в передаче нерв ного импульса следующему нейрону определяется закономерным выбросом путем экзоцитоза в синаптическую щель медиатора.
Рис. 172. Схема ультрамикроскопического строения различных типов си напсов:
А — питотопография синапсов; В — синапс тормозного типа; В — синапс возбуди тельного типа; Г — электротонический синапс; 1 — аксосоматический синапс; 2— аксодендритические синапсы; з — аксоаксональный синапс; 4 — дендриты; 5 — дендритный шипик; 6 — аксон; 7 — синаптичеекие пузырьки; 8 — пресинаптическая мембрана; 9 —• постсинаптическая мембрана; 10 — синаптическая щель; и— постсинаптические структуры.
21?
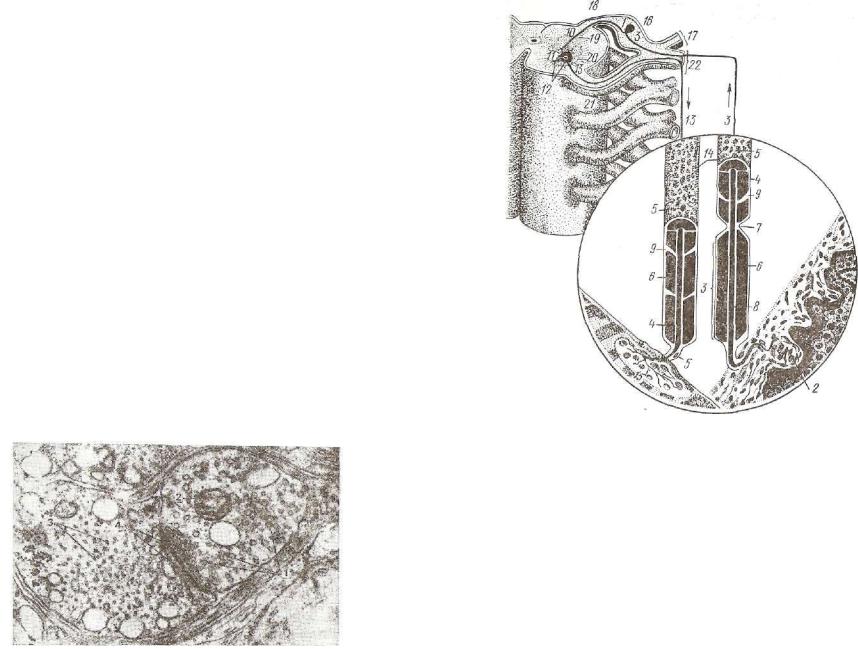
"Последний, поступая на мембрану постсинаптического полюса,, вызывает изменение ее проницаемости, волну деполяризации — генерацию нервного импульса. Роль медиатора, кроме указанных выше, могут играть и другие вещества, а именно: адреналин, се* ротонин, гамма-аминомасляная кислота и др.
Синаптические пузырьки, несущие различные медиаторы,, морфологически различны. В холинэргических синапсах они мелкие (30—40 нм) и прозрачные. В их составе иногда содержит ся также несколько очень крупных и электроноплотных пузырь ков (80—150 нм), химический состав и значение последних в= настоящее время неясны. Предположительно считают, что опт содержат биогенные амины. Синаптические пузырьки адренэргических синапсов крупнее (50—90 нм) и морфологически харак теризуются наличием в них электроноплотной гранулы. Выделе ние медиатора происходит экзоцитозом в синаптическую щель содержимого синаптических пузырьков.
П о с т с и н а п т и ч е с к а я м е м б р а н а холинэргических си напсов содержит «холинрецептивный» белок. При взаимодейст вии с ацетилхолином происходят конформационные изменения его молекул, приводящие к изменению проницаемости мембран и генерации нервного импульса в нейроне (рис. 173). Медиатор тормозных синапсов — гамма-аминомасляная кислота не увеличи вает проницаемость постсинаптической мембраны для ионов, а уменьшает ее и, следовательно, стабилизирует мембранный потен циал, то есть тормозит генерацию нервного импульса.
На синаптических мембранах имеются характерные специа лизации. Под мембранами как пресинаптического, так и пос гсинаптического полюсов отмечают скопления плотного материала и тонких филаментов. Скопления обычно толще на пресинаити-
Рис. 173. Электронная микрофотография аксодендригического синапса в краниальном шейном узле кота (препарат Козлова):
I — синаптические пузырьки; 2 — митохондрия; з — нейротрубочки в цитоплазме дендрита; 4 — утолщенная постсинаптическая мембрана.
1 — чувствительная нервная клетка; 2 — рецептор в коже; 3 — дендрит чувстви тельной клетки; 4 — оболочка; б — ядро леммоцита; б — миелиновая оболочка; 7 — перехват нервного волокна; 8 — осевой цилиндр; 9 — насечка на нервном во локне; 10 — нейрит чувствительной клетки; и — двигательная клетка; 12 — ден дрит двигательной клетки; 13 — нейрит двигательной клетки; И — миелиновыв- •волокна; 15 — эффектор (моторная бляшка); 16 — спинномозговой узел; П — дор сальная ветвь спинномозгового нерва; IS — задний корешок; 19 — задний рог; 20— передний рог; 21 — передний корешок; 22 — вентральная ветвь спинномозгового яерва.
ческой мембране. С пресинаптическими уплотнениями часто свя заны синаптические пузырьки. На тангенциальных срезах уп лотнений видно, что они не гомогенны, а состоят из шестиуголь ных и треугольных структур, в центре которых можно различить синаптический пузырек. Скопления пузырьков и уплотнения вме сте называют с и н а п т и ч е с к и м к о м п л е к с о м , а поскольку о пи, по-видимому, являются местами преимущественного скопле
ния пузырьков и выделения |
медиатора, их называют |
также |
||
а к т и в н ы м и з о н а м и . |
В |
области синапсов |
выявляют |
и ма |
ленькие прикрепительные |
устройства — точки |
п р и к р е п л е - |
||
219
218
ния (punctum adherens). От уплотнений синаптических комплек сов они отличаются большей толщиной и симметричностью и ма лой линейной протяженностью.
Электротоиические синапсы образуются при плотном приле жании плазмолемм, двух нейронов, преимущественно их дендритов, и перикариона.
Нервная система организма представлена чувствительными, ассоциативными и двигательными клетками, объединенными межнейрональными синапсами в функционально активные образова ния — р е ф л е к т о р н ы е дуги . Простая рефлекторная дуга со
стоит |
из двух нейронов — чувствительного и двигательного |
(рис. |
174). |
В подавляющем большинстве рефлекторные дуги высших по звоночных животных содержат в своем составе еще значитель ное количество ассоциативных нейронов, расположенных между чувствительным и двигательным нейронами.
РАЗДЕЛ ШУЗ
Г Л А В А 4
ОРГАНЫ НЕРВНОЙ СИСТЕМУ
Деление нервной системы на центральную, к которой относят головной и спинной мозг, и периферическую — нервные стволы
инервные окончания условно и допустимо лишь из методических соображений. Морфологическим субстратом рефлекторной дея тельности нервной системы являются рефлекторные дуги, объ единяющие цепь нейронов, различные звенья которой лежат как в периферических нервных узлах, так и в сером веществе цен тральной нервной системы. Функционально всю нервную систему делят на соматическую (или цереброспинальную), иннервирующую тело животного, кроме внутренних органов, сосудов и желез,
ивегетативную, регулирующую деятельность последних.
Развитие нервной системы. Органы нервной системы разви ваются из нервной трубки и ганглиозной пластинки. Из краниаль ной части нервной трубки дифференцируются головной мозг и органы чувств. Из ее туловищного отдела и ганглиозной пластин ки образуются спинной мозг, спинальные и вегетативные ганглии и хромаффинная ткань организма. Интенсивно развиваясь, бо ковые отделы нервной трубки формируют дорсальную — крыльную пластинку и вентральную — основную. На этой стадии разви тия в боковых стенках нервной трубки различают три зоны: внут
реннюю, выстилающую |
спинномозговой канал, — э п е н д и м у , |
||
п л а щ е в о й |
с л о й , |
из которого в дальнейшем развивается серое |
|
вещество, и |
к р а е в |
у ю |
в у а л ь , формирующую белое вещества |
спинного мозга. Нейробласты вентральной зоны плащевого слоя— передних столбов — дифференцируются в двигательные нейроны ядер вентральных рогов. Их аксоны, выходя из спинного мозга,, образуют вентральные корешки. В дорсальных столбах и проме жуточной зоне формируются ядра вставочных (ассоциативных) клеток. Их аксоны в белом веществе образуют его проводящие пути. В дорсальные рога поступают нейриты чувствительных клеток спинальных ганглиев.
Спинальные, периферические вегетативные узлы и хромаф финная ткань развиваются из ганглиозной пластинки.
СПИННОМОЗГОВЫЕ УЗЛЫ
В процессе эмбриогенеза ганглиозная пластинка расчленяется на отдельные нервные узлы, на поверхности которых формиру ется соединительнотканная оболочка. Клетки ганглиозной пла-
221
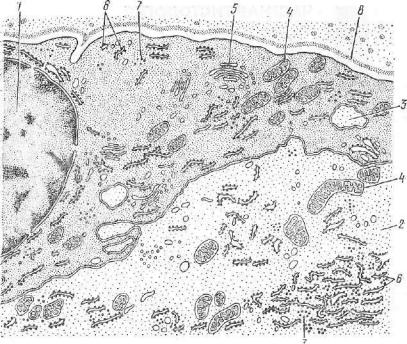
I — ядро клетки сателлита; 2 — цитоплазма нейрона; в — выпячивания цитоплазмы, внедряющиеся в клетку-сателлит; 4—митохондрии; 5—комплекс Гольджи- 6—ше роховатая эндоплазматическая сеть; 7 — свободные рибосомы; 8 — базальная мем-
стинкп в процессе эмбриогенеза развиваются в двух направлениях. Одни из них превращаются в нейробласты и соответственно диф ференцируются в характерные для чувствительных нервных кле ток биполярные нейроциты, содержащие два отростка — дендрит и нейрит. У высших позвоночных животных эти отростки, сбли жаясь, формируют в своем начальном отделе единый тяж цито плазмы (за что эти клетки получили название псевдоуниполяр ных), который на некотором расстоянии Т-образно делится на „два отростка — дендрит, идущий на периферию в составе сме шанного нерва в качестве его чувствительного волокна, и ней рит, в^ комплексе с нейритами других нейроцитов узла обра зующий дорсальный корешок спинного мозга и поступающий в последний.
Нейроциты спинальных ганглиев покрыты одним слоем кле ток глии — сателлитов нервных клеток. Наружный слой капсулы нейрона образован тонковолокнистой соединительнотканной кап сулой. Клетки последней в отличие от сателлитов характеризу ются овальной формой ядер (рис. 175).
•222
Анатомически спинной мозг состоит из двух симметричных по ловинок, отграниченных друг от друга вентральной срединной щелью и дорсальной срединной перегородкой. Центрально рас
положенное |
с е р о е в е щ е с т в о спинного мозга содержит муль- |
типолярпые |
нервные клетки, образующие ядра спинного мозга. |
Ifериферическая часть его — белое в е щ е с т в о — представле на совокупностью нервных волокон, входящих в состав различ ных проводящих путей центральной нервной системы.
Серое вещество спшшого мозга. Анатомически серое вещест во спинного мозга состоит из двух половин, связанных компссурой. Каждая из них имеет д о р с а л ь н ы е и в е н т р а л ь н ы е р о г а. В грудных и поясничных сегментах спинного мозга можновыделить верхнебоковой отдел вентральных рогов как его лате ральные рога. В серой комиссуре, связывающей две половины серого вещества, находится центральный канал спинного мозга.
Серое вещество образовано мультиполярными нейронами,, безмиелиновыми и миелиновыми нервными волокпами и нейроглией. Группы нервных клеток одинакового функционального» значения образуют ядра серого вещества.
По морфологическим признакам, локализации, участию в нерв ном проведении в составе серого вещества спинного мозга мож но выделить следующие виды клеток: к о р е ш к о в ы е клет ки — клетки, нейриты которых покидают спинной мозг в составе
его вентральных корешков, в н у т р е н н и е к л е т к и , |
нейриты |
||
которых образуют синапсы на клетках серого вещества |
спинного |
||
мозга, и |
п у ч к о в ы е к л е т к и . Их нейриты формируют в белом |
||
веществе |
обособленные пучки, проводящие нервные импульсы от |
||
определенных ядер спинного мозга в его другие |
сегменты или в- |
||
определенные отделы головного мозга, формируя |
проводящие пу |
||
ти центральной нервной системы. |
|
|
|
В сером веществе спинного мозга выделяют участки, отли чающиеся по нейрональному составу, характеру нервных воло кон и нейроглии. Так, в дорсальных рогах серого вещества сле дует выделить губчатый слой, желатинозное вещество, собствен ное ядро дорсального рога, его дорсальное ядро, или ядро Кларка.
Губчатый слой д о р с а л ь н ы х |
р о г о в серого |
вещества со |
|
держит мелкие пучковые клетки, погруженные |
в |
широкопетли |
|
стый глиальный остов. |
|
|
|
Ж е л а т и н о з н о е в е щ е с т в о |
образовано |
преимуществен |
|
но элементами глии, в которой содержатся в незначительном ко личестве мелкие пучковые клетки.
В составе дорсального рога располагаются собственное ядро заднего рога, грудное ядро (ядро Кларка) и значительное ко личество диффуэио рассеянных мелких мультиполярных вста вочных нейронов.
С о б с т в е н н о е я д р о дорсального рога содержит пучковыеклетки, аксоны которых через переднюю белую спайку переходят
22$