

gistologia
.pdfторых формах анемий центральная бледно окрашенная часть эри троцитов увеличена — гипохромные эритроциты. При суправитальном окрашивании капли крови бриллиантовым крезиловым синим в приготовленном затем мазке можно обнаружить молодые формы эритроцитов, содержащие зернисто-сетчатые структуры. Такие клетки называют ретикулоцитами, они являются непосред ственными предшественниками эрелых эритроцитов. При элек тронной микроскопии в содержимом ретикулоцитов обнаружены компоненты белоксинтезирующей системы — рибосомы, элементы эндоплазматической сети, митохондрии. Радиоавтографическим методом доказано, что в ретикулоцитах продолжает осуществлять ся синтез белка гемоглобина. Подсчет ретикулоцитов использует ся для получения информации о скорости образования эритроци тов. Обнаружение в большем количестве по сравнению с нормой ретикулоцитов в крови — признак усиленного эритропоэза.
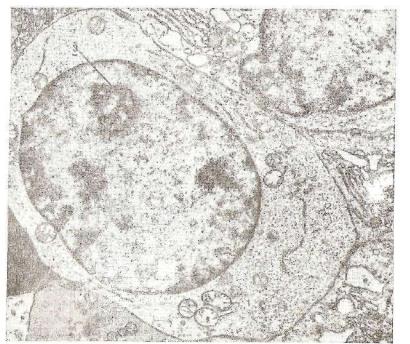
В зрелых эритроцитах при электронной микроскопии не удает ся выявить никаких органелл, их внутреннее содержимое имеет высокую электронную плотность.
В период между 100—130 днями |
(у кроликов |
через 45— |
60 дпей) после выхода из красного |
костного мозга |
эритроциты |
фагоцитируются макрофагами главным образом в селезенке, пече ни и красном костном мозге. При этом от гема гемоглобина от щепляется железо, а из оставшейся части молекулы образуются пигменты желчи, мочи и кала. Железо с помощью белка плазмы крови транспортируется главным образом в костный мозг, где вновь используется при синтезе гемоглобина развивающимися эритроцитами. Таким образом, железо, освобождающееся из фа гоцитированных эритроцитов, почти полностью доступно для реутилизации. Избыток железа аккумулируется в макрофагах селезенки или печени в виде гранул гемосидерина, которые выяв ляют гистохимическими методами.
Эритроциты обладают свойством противостоять различным разрушительным воздействиям — осмотическим, механическим и др0 При значительных изменениях концентрации солей в окружа ющей среде, например при помещении крови в гипотонический раствор, эритроциты набухают, приобретают сферическую форму; мембрана перестает удерживать гемоглобин, и он выходит в окру жающую жидкость — явление г е м о л и з а . Выход гемоглобина из эритроцитов может происходить в организме при действии зме иного яда, токсинов, выделяемых некоторыми бактериями, возбу дителями паразитарных болезней. Гемолиз развивается также при переливании несовместимой по группе крови. Практически важ но при введении в кровь животным жидкостей осуществлять кон троль ва тем, чтобы вводимый раствор был изотоническим.
У эритроцитов по сравнению с плазмой и лейкоцитами крови относительно большая плотность (удельный вес). Если кровь, предварительно обработанную противосвертывающими вещества ми, поместить в какой-либо сосуд, то отмечают оседание эритро цитов. С к о р о с т ь о с е д а н и я э р и т р о ц и т о в (СОЭ) у жи-
124
вотных разного возраста, пола и вида неодинакова. Высокая СОЭ у лошадей и, наоборот, низкая у крупного рогатого скота. Измене ния СОЭ, наблюдаемые в патологии, имеют диагностическое и прогностическое значение.
Лейкоциты
Лейкоциты — разнообразные по морфологическим признакам и функциям клетки сосудистой крови. В организме животных они выполняют многообразные функции, направленные прежде всего на защиту организма от чужеродного влияния путем фагоцитар ной активности, участия в формировании гуморального п клеточ ного иммунитета, а также в восстановительных процессах при тканевом повреждении.
В крови животных лейкоцитов в 600—800 раз меньше, чем эритроцитов. В 1 мкл крови у крупного рогатого скота их насчи тывают 4,5—12,0 тыс., у лошадей —7,0—12,0 тыс., овец —6,0— 14,0 тыс., свиней — 8,0—16,0 тыс., собак — 8,5—10,5 тыс., кур — 20,0—40,0 тыс. Увеличение количества лейкоцитов — л е й к о ц и те з — характерный признак для многих патологических процес сов, но может наблюдаться и у здоровых животных (лейкоцитозноворожденных, пищеварительный лейкоцитоз и т. д.).
Образовавшись в кроветворных органах и поступив в кровь,, лейкоциты лишь непродолжительное время пребывают в сосуди стом русле, затем они мигрируют в вокругсосудистую соединитель ную ткань и органы, где осуществляют свою основную функцию. Для многих лейкоцитов тканевая фаза их жизни является завер шающей.
Особенность у лейкоцитов та, что они обладают подвижностью; В цитоплазме лейкоцитов, непосредственно около плазмолеммы, имеется комплекс актиновых микрофиламентов, с помощью кото рых приводится в движение клеточная поверхность и образуются псевдоподии. Изменяя свою внешнюю форму и форму ядра, белые кровяные клетки способны активно перемещаться между клетка ми эндотелия сосудов, проникать через базальные мембраны и клетки эпителия, мигрировать в основном веществе соединитель ной ткани.
В лейкоцитах различают ядро и цитоплазму, содержащую раз личные органеллы и включения. Классификация лейкоцитов ос нована на учете их морфологических признаков, выявляемых при световой микроскопии окрашенных мазков крови и имеет прежде всего клинико-практическое значение. Те лейкоциты, в цитоплаз ме которых содержится специфическая зернистость, называются з е р н и с т ы м и (или гранулоцитами). Зрелые зернистые лейко циты, как правило, имеют расчлененное на сегменты ядро — сегментоядерные клетки. В соответствии с различием в окрашивании цитоплазматической зернистости в группе гранулоцитов выделя ют три вида клеток: н е й т р о ф и л ы — зернистость окрашивается и кислыми, и основными красителями; э о з и н о ф и л ы — вер-
125.

иистость |
окрашивается |
кислыми красителями и |
б а з о ф и л ы |
— |
||||
зернистость окрашивается основными |
красителями. Н е з е р н и с |
|||||||
тые |
л е й к о ц и т ы |
(агранулоциты) |
характеризуются отсутст |
|||||
вием специфической зернистости в цитоплазме |
и несегментиро- |
|||||||
занными |
ядрами. В |
группе |
агранулоцитов |
выделяют |
два |
|||
вида |
— л и м ф о ц и т ы |
и |
м о н о ц и т ы . Таким образом, схема |
|||||
классификации лейкоцитов |
выглядит |
следующим образом: |
|
|||||
|
|
|
|
о„„„ |
„„ттл |
|
Нейтрофилы |
|
-(iSSSSSL) ' >Эози|о1ияы
Лейкоциты |
|
ь |
lP ^ |
' |
Базофилы |
|
|
||||||
|
|
|
|
Незернистые |
|
., Лимфоциты |
|
|
|
(агранулоциты) |
Моноциты |
||
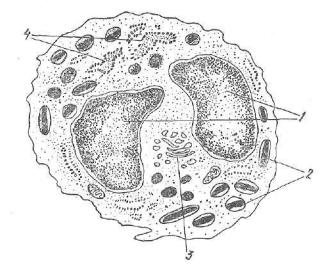
В ветеринарной практике при анализе крови животных важ ное диагностическое значение имеет дифференциальный подсчет лейкоцитов. Определенное процентное соотношение между отдель ными видами лейкоцитов называют лейкоцитарной формулой —• лейкограммой. При определении лейкоцитов на окрашенных маз ках крови с помощью светового микроскопа учитывают многие морфологические признаки: величину клеток, форму и плотность ядра, соотношение между объемом ядра и цитоплазмы, наличие или отсутствие в цитоплазме зернистости, ее окраску, величину и распределение в объеме цитоплазмы (рис. 96). В настоящее вре мя лейкоцитарную формулу устанавливают с помощью автомати ческих приборов.
Г р а н у л оциты (зернистые лейкоциты).
Нейтрофильные лейкоциты (нейтрофилы) — очень подвижные клетки, обладающие высокой фагоцитарной активностью.
1 — эритроциты; 2 — нейтрофилы; 3 — эозинофил; 4 — базофил; 5 — малый лим фоцит; б — средний лимфоцит; 7 — моноциты,
126
2. Лейкограмма крови животных,
0—2 |
5—8 |
0 - 1 |
2—5 |
20—30 |
40—65 |
2—7 |
|
0—1 |
2—6 |
0—1 |
3-6 |
45-65 |
25—44 |
2—4 |
|
0—1 |
4—12 |
0—2 |
3—6 |
35—45 |
40—50 |
2—5 |
|
0—1 |
1—4 |
0—2 |
2—4 |
40—48 |
40—50 |
2—6 |
|
0—1 |
3—9 |
0 |
1—6 |
45—70 |
20-40 |
1-5- |
|
0—1 |
3—7 |
0 |
2—5 |
55—65 |
20-35 |
|
1-4 |
0—1 |
4—12 |
0 - 2 |
1—6 |
40—52 |
30—45 |
1-5 |
|
0—2 |
4—12 |
0 |
1—5 |
30—45 |
35—54 |
3—8 |
|
0—1 |
1—5 |
0 |
1—4 |
20—35 |
55—75 |
1-5 |
|
Крупный рогатый скот Лошади Овцы Свиньи Собаки
Северные олени Верблюды Морские свинки Крысы белые
И. И. Мечников назвал их м и к р о ф а г а м и . Нейтрофилы всего лишь несколько часов циркулируют в сосудистой крови. Благо даря х е м о т а к с и с у — направленному движению фагоцитов под влиянием хемотаксических факторов нейтрофилы мигрируют из кровеносных капилляров в соединительную ткань, накаплива ются в очаге воспаления, где и осуществляют свою основную фа гоцитарную функцию, обеспечивая очищение очага воспаления от микроорганизмов и продуктов клеточного и тканевого распада. В процессе фагоцитоза нейтрофилы погибают и вместе с бактери альными веществами и остатками разрушенных тканей образуют массу, называемую гноем. Хемотаксическими свойствами облада ют многие продукты экзо- и эндогенного происхождения и преж де всего вещества, выделяемые бактериями и вирусами, самими лейкоцитами, а также образующиеся при тканевом распаде.
В крови животных нейтрофилов содержится от 25 до 70% всех лейкоцитов. Больше всего нейтрофилов у лошадей, северных оленей, собак — до 65—70%. Это животные с так называемым нейтрофильным профилем крови. Н е й т р о ф и л е з — увеличение в лейкоцитарной формуле процента нейтрофилов — наиболее ти пичен для гнойно-воспалительных процессов.
В мазках крови, окрашенных по Романовскому—Гимзе, раз мер нейтрофилов от 8 до 15 мкм, их определяют главным образом по форме и структуре ядра. У зрелых нейтрофилов ядро состоит из нескольких сегментов, соединенных тонкими перемычками. Очень большая сегментированность ядер характерна для нейтро филов крови овец. У этих животных часто встречаются клетки с 8—10 сегментами. В сегментах содержится плотный примембранный хроматин, благодаря чему они интенсивно окрашиваются.
В крови животных (см. лейкограмму) имеется определенное ко
127
личество менее зрелых предшественников сегментированных
форм — палочкоядерных нейтрофилов, имеющих ядро в виде изо
гнутой палочки или буквы S и метамиелоцитов (юных), содержа имеетклиникещихбобовидноопределенно. Пр многихядрое.диагностическоСоопа ношениологическиэтиххзначениесостоянияхтрехвидов ветуменьшаетсянейтрофиловринарнй
•количество сегментоядерных и нарастает количество палочкоядер- •ных и юных нейтрофилов, мобилизуемых из красного костного мозга в сосудистую кровь. Обогащение крови более молодыми формами нейтрофилов получило название «сдвига влево», так как при выведении лейкограммы незрелые клетки принято записы вать слева, а зрелые — справа. Резкий сдвиг влево наблюдают при тяжелых септических инфекциях.
Цитоплазма нейтрофилов слабооксифильна, в ней содержится очень мелкая, трудно различимая при световой микроскопии -окрашенных мазков крови зернистость. Отношение зернистости к красителям варьирует у разных животных: у собаки, кошкп и -свиньи зернистость окрашивается в розово-фиолетовый цвет. •У кроликов гранулы окрашиваются эозином в ярко-красный цвет, то есть имеют выраженное сродство к кислым красителям. По этой причине нейтрофилы кроликов иногда называют псевдоэозинофилами. Оксифильна и палочковидная зернистость нейтрофилов у птиц.
При электронно-микроскопическом исследовании в цитоплазме нейтрофилов обнаруживают небольшое количество органелл об щего назначения, включения гликогена, в периферической зоне •содержатся микрофиламенты, обеспечивающие движение клетки. Характерным для цитоплазмы нейтрофилов является наличие в ней гранул двух разновидностей: азурофильных и специфических, содержимое которых позволяет этим клеткам выполнять свои •функции. Появляющиеся на ранних стадиях развития более круп ные и электроноплотные азурофильные гранулы содержат, кроме типичных гидролитических ферментов, миелопероксидазу и лизоцим. В многочисленных (до 80% всего количества гранул), но бо лее мелких специфических гранулах выявляется щелочная фосфатаза. Показано, что с фагосомой первой по времени контактиру ет и сливается специфическая гранула, в последующем переваривание осуществляется с помощью ферментов азурофиль ных гранул. Дегрануляция нейтрофилов и активизация лизосомальных гидролитических ферментов происходит и при разруше нии клеток, что наблюдается при воспалительной реакции. Таким образом, благодаря совместному действию многочисленных фер ментов, содержащихся в гранулах, нейтрофилы могут перевари вать все макромолекулы, присутствующие в бактериях и очаге воспаления,
Эозинофильиые лейкоциты (эозинофилы) — разновидность гранулоцитов. Количество их у животных составляет 2—12% всех лейкоцитов сосудистой крови. Наибольший процент эозинофилов в крови крупного рогатого скота, верблюдов и овец. По размеру •эозинофилы крупнее нейтрофилов и имеют диаметр 12—18 мкм. Ядро в эрелых эозинофилах также сегментированное, но чаще ядро состоит из двух сегментов. Значительную сегментированность ядра (3—5 сегментов) отмечают в эозинофилах овец. В кро- •ви животных встречаются палочковидные и юные формы эозино филов.
3
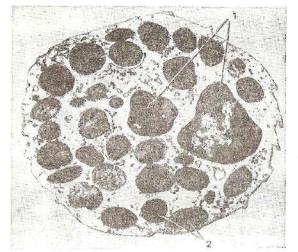
Рис. 97. Схема ультрамикроскопического строения эозино-
фюпьного гранулоцита крысы:
2 — сегменты ядра; 2 — зрелые специфические гранулы, со держащие кристаллоиды; з — комплекс Гольджи; 4 — грану лярная эядоплазматическая сеть (рис. Козлова).
Весьма характерна для эозинофилов относительно крупная специфическая зернистость цитоплазмы. Гранулы, как правило, равномерно и плотно заполняют площадь цитоплазмы. Зерна ярко окрашиваются в оранжево-красный или красный цвет, то есть имеют выраженную оксифилию вследствие содержания в них основных белков. Особенно крупные размеры гранул в эозинофи лах однокопытных (лошадь, осел). Гистохимическими методами в зернистости выявлены различные ферменты: кислая фосфатаза, пероксидаза, гистаминаза, арилсульфатаза и др.
При электронной микроскопии зрелые гранулы эозинофилов некоторых видов животных имеют расположенные в центре элек троноплотные палочковидные структуры, окруженные по пери ферии более светлым содержимым (рис. 97).
Подобно нейтрофилам, эозинофилы обладают хемотаксисом. Хемотаксическими веществами в отношении эозинофилов являют ся комплексы антиген—антитело, гистамин и другие низкомоле кулярные факторы, выделяемые тканевыми базофилами. По срав нению с нейтрофилами эозинофилы менее подвижны и обладают меньшей фагоцитарной активностью. Во время фагоцитоза проис ходит дегрануляция эозинофилов, высвобождение из гранул мно гих ферментов, с помощью которых осуществляется инактивация гистамина и других медиаторов воспаления. Таким образом, важ ная функция эозинофилов — антигистаминовая, участие в ограни чении воспалительного процесса.
Эозинофилы являются важнейшими эффекторными клетками в противопаразитарном иммунитете. Многие паразитарные болез-
«28 |
9 Заказ № 90S |
129 |
|
ни животных (фасциолез, эхинококкоз и др.) сопровождаются усилением продукции эозинофилов в красном костном мозге и уве личением их количества в сосудистой крови (у крупного рогатого скота до 20%). В антипаразитарном механизме важнейшее зна чение имеет прямой контакт эозинофилов с возбудителем, поэто му они в больших количествах скапливаются вокруг внедривших ся в организм паразитов. Характерно содержание значительного количества эозинофилов в рыхлой соединительной ткани стенки кишечника.
Базофильные лейкоциты — базофилы — самая малочисленная разновидность гранулоцитов периферической крови животных (0,5—2% всех лейкоцитов). В крови сельскохозяйственных птиц 3—4% базофалов, а содержание их в крови земноводных доходит до 20 %. В мазках окрашенной крови диаметр базофилов несколь ко меньший, чем у других гранулоцитов, — 10—12 мкм. Ядро ча ще в виде неправильно изогнутой палочки, реже сегментирован ное. Слабооксифильная цитоплазма содержит неравномерно распределенную, различную по величине и форме, легко раствори мую в воде зернистость. При окраске по Романовскому—Гимз© зернистость красно-вишневого цвета, то есть обладает метахромазией — окрашивается в цвет, отличающийся от цвета основного красителя (азура). Это свойство метахромазии обусловлено особой формой полимеризации молекул красителя при его взаимодейст вии с гепарином, входящим в состав гранул. Кроме гепарина, в базофилах имеются гранулы, содеря^ащие высокоактивное веще ство — гистамин. Электронно-микроскопически гранулы неодина ковы по электронной плотности (рис. 98).
На поверхности базофилов расположены специальные рецеп торы, с помощью которых связываются иммуноглобулины Е. При соединение антигенов (аллергенов) и образование комплекса антиген—антитело вызывают дегрануляцию базофилов и выход значительного количества содержащихся в них фармакологически активных веществ — гистамина и гепарина. Быстрое высвобож дение этих соединений и лежит в основе реакций повышенной чувствительности немедленного типа — сывороточной болезни, фе номена Артюса и др. Установлено, что базофилы составляют зна чительную часть клеточного инфильтрата при различных формах гиперчувствительности замедленного типа. Таким образом, базо филы участвуют в иммунологических реакциях организма, в част ности в реакциях аллергического типа.
А г р а и у л о ц и т ы (незернистые лейкоциты). Моноциты, В 1 мкл крови у животных содержится от 1 до 8%. Моноциты периферической крови — самые крупные из лейкоцитов — в маз ках окрашенной крови их диаметр 18—20 мкм и более. Это клетки с относительно большим количеством голубовато-серой цитоплаз мы (с мелкой азурофильной зернистостью) и различным по сво ей форме ядром. Последнее чаще всего бобовидной или лопастной формы, в нем различают расположенный преимущественно по периферии конденсированный хроматин и мелкие ядрышки. При
Рис. 98. Электронная микрограмма базофильного гранулоцнта:
1 — сегменты ядра; г — базофильные гранулы (по Блюму и Фаусету).
электронной микроскопии в цитоплазме выявляются лизосомы, пиноцитозные пузырьки, развитый комплекс Гольджи, небольшие по размеру митохондрии и плотные гранулы, соответствующие азурофильным гранулам, видимым в световой микроскоп (рис.99). Цитохимическими методами в моноцитах обнаруживают липиды, гликоген; в них высокая активность неспецифической эстеразы, имеются кислые гидролазы, пероксидаза, лизоцим (мурамидаза), с участием которых обеспечивается антимикробное действие. По цитохимической характеристике моноциты имеют сходство с клет ками нейтрофильного ряда.
Циркулирующие в крови моноциты являются предшественни ками тканевых и органных макрофагов, на основании чего они отнесены к системе мононуклеарных фагоцитов (СМФ). После пребывания в сосудистой крови (12—36 ч) моноциты мигрируют через эндотелий капилляров и венул в ткани и превращаются в фиксированные и подвижные макрофаги. При этом в них отмеча ется сильное развитие лизосом и появление фагосом. В большом количество моноциты выходят из кровяного русла в ткани при воспалительных реакциях, в том числе и иммунных. В очагах хронического воспаления моноциты становятся важнейшим кле точным элементом, выполняющим многообразные функции (сти мулируют фибробластические процессы, образуют многоядерные клетки и др.).
Лимфоциты — важнейшие клетки, участвующие в разнообраз ных иммунологических реакциях организма. Это многочисленная группа лейкоцитов крови животных. У крупного рогатого скота,
130 |
9* |
I3L |
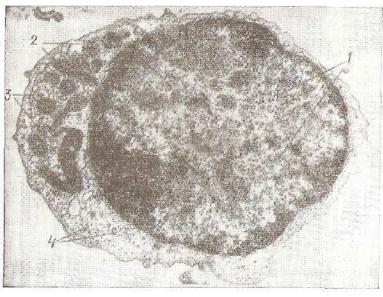
Рис. 99. Электронная микрограмма среднего лимфоцита:
1 — ядро; 2 — митохондрии; 3 — мелкие канальцы гранулярной эндоплазматической сети; 4 — свободные полисомы.
овец, кроликов, белых крыс число лимфоцитов составляет 40— 65% всех лейкоцитов. У других животных (лошади, собаки, свиньи, северные олени), отличающихся преобладанием в крови нейтрофилов, лимфоцитов содержится 20—40%. Большое количе ство лимфоцитов находится в лимфе.
По размеру и некоторым структурным особенностям, выявляе мым при световой микроскопии окрашенных мазков крови, разли чают малые (диаметр до 8 мкм), средние (8—11 мкм) и большие (более 11 мкм) лимфоциты. Малые лимфоциты составляют ос новную массу (до 90%) всех лимфоцитов крови животных. Это клетки с выраженным преобладанием объема ядра над объемом цитоплазмы. Ядро у них чаще всего круглое с небольшим углуб лением. Крупные конгломераты конденсированного хроматина расположены как по периферии, так и в центральной части ядра, что делает его темным и маскирует ядрышко. Базофильная цито плазма обнаруживается или в виде узкого вокругядерного ободка, или в виде серпа, прилежащего к одной стороне ядра. В цитоплаз ме некоторых лимфоцитов содержится небольшое количество мел ких азурофильных гранул.
Для средних и больших лимфоцитов более характерны ядра слегка бобовидной формы с меньшим количеством конденсирован ного хроматина и отчетливо заметным ядрышком, а также более
широкий ободок цитоплазмы. Электронно-микроскопически в цитоплазме малого лимфоцита выявляют свободные полисомы и рибосомы. Другие органеллы — митохондрии, мелкие канальца эндоплазматической сети, элементы комплекса Гольджи, как пра вило, немногочисленны и располагаются преимущественно наряду с центриолями около ядерного углубления.
Морфологически сходные малые лимфоциты крови являются функционально разнородными клетками, развитие которых до их поступления в сосудистую кровь происходит в различных орга нах.
Различают два основных класса лимфоцитов: Т- и В-лимфо- циты.
Т - л и м ф о ц и т ы развиваются из костномозговых клеток в корковой части долек тимуса. Их дифференцировка выражается в появлении в плазмолемме поверхностных антигенных маркеров (антигены гистосовместимости) и многочисленных рецепторов, с помощью которых происходит распознавание чужеродных анти генов и иммунных комплексов.
В-л и м ф о ц и т ы свое название получили потому, что у птиц этот вид лимфоцитов образуется из стволовых предшественников в фабрициевой сумке (Bursa — В). Местом развития В-лимфоци- тов у животных считают миелоидную ткань костного мозга. В ходе дифференцировки В-лимфоциты приобретают способность синтези ровать иммуноглобулины, которые становятся составными элемен тами их плазмолеммы и выполняют роль рецепторов. С их участи ем В-лимфоциты взаимодействуют с антигенами при их распозна вании. Кроме того, на поверхности В-лимфоцитов образуются рецепторы для связывания комплемента (его компонента Сз) и рецепторы для Fc фрагмента иммуноглобулиновых молекул.
Установлено, что в основном (более 70%) малые лимфоциты сосудистой крови отвносятся к Т-лимфоцитам, среди которых зна чительное количество долгоживущих клеток памяти (10 лет и более). Большинство В-лимфоцитов относится к короткоживущим (недели—месяцы).
Циркулирующие в крови Т- и В-лимфоциты представляют под вижную популяцию относительно функционально неактивных клеток. Вне сосудистого русла они скапливаются в определенных участках периферических лимфоидиых органов (для Т-лимфоци- тов это тимуезависимые ЗОБЫ) , где могут при действии соответ ствующих антигенов подвергаться активации, превращаться в бластпьто формы, размножаться и дифференцироваться в эффекторные клетки клеточного и гуморального иммунитета, а также создавать увеличенный фонд клеток памяти.
Эффекториыми клетками в системе Т-лимфоцитов являются три основные субпопуляции: Т-киллеры (цитотоксические лимфо циты), Т-хелперы (помощники) и Т-супреесоры (угнетающие). Эффекторными клетками В-лимфоцитов являются плазмобласты и зрелые плазмоциты, способные в повышенном количестве про дуцировать иммуноглобулины.
132 |
133 |
Субпопуляции Т-системы также отличаются своими поверхно стными маркерами, биологическими свойствами и выполняют раз личные специфические иммунологические функции, которые со ставляют основу двух различных иммунных ответов — клеточного и гуморального.
Цитотоксические лимфоциты (Т-киллеры — убийцы) путем непосредственного контактного воздействия или с помощью близ кодействующих токсических медиаторов (лимфокинов) разрушают чужеродные клетки-мишени или изменившиеся собственные клет ки. Такой тип реакции и обезвреживания антигенов (не сопро вождающийся выработкой растворимых антител) называют кле точным иммунитетом. Он наблюдается при отторжении трансплан татов, при реакциях гиперчувствительности замедленного типа, при разрушении опухолевых клеток и развитии аутоиммунных реакций.
Т-хелперы и Т-супрессоры — две субпопуляции клеток, имею щих значительную роль в регуляции активности В-лимфоцитов. Т-хелперы выделяют продукты своей синтетической деятельности (медиаторы), которые, взаимодействуя с В-лимфоцитами, способ ствуют их превращению в антителопродуцирующие плазматиче ские клетки. Т-супрессоры подавляют процессы деления В-клеток и образования плазмоцитов и таким образом оказывают тормозя щий эффект на антителообразование. Согласпо современным пред ставлениям, в механизме действия клеток хелперов и супрессоров важнейшее значение отводится Fc рецепторам, находящимся на поверхности В-клеток. Вспомогательный эффект Т-хелперов рас сматривают как блокировку этих Fc рецепторов медиаторами хел перов, что препятствует связыванию рецепторов иммуноглобули нами, но способствует их связыванию со специфическими антиге нами. Медиаторы Т-супрессоров способствуют образованию связи между иммуноглобулинами и Fc рецепторами В-клеток, тем самым ингибируют их превращение в плазмоциты. В тех случаях, когда антиген вызывает в организме образование антител, В-лимфоциты находятся под влиянием Т-хелперов и макрофагов. Показано, что для развития гуморального иммунного ответа на тимусзависимые антигены превращение В-лимфоцита в плазмоцит не может про исходить без участия всех трех типов клеток: Т-хелперов, В-лим фоцитов и макрофагов. Имеется ряд гипотез о том, каким обра зом происходит взаимодействие клеток в процессе этого иммунного ответа.
На часть тимуснезависимых антигенов (пневмококковый поли сахарид, липополисахарид кишечной палочки и др.) В-лимфоци ты отвечают самостоятельно. Этим антигенам свойственна особая структура молекул, позволяющая им концентрироваться на по верхности В-клеток без помощи Т-хелперов.
Существенная |
функция Т-лимфоцитов — их способность |
син |
|||
тезировать |
и выделять ряд факторов белковой природы — |
лим |
|||
ф о к и н о в , |
которые изменяют |
поведение |
других клеток |
(фак |
|
тор, угнетающий |
миграцию |
макрофагов, |
фактор активации |
||
макрофагов, хемотаксические факторы для нейтрофилов. эозинофилов, базофилов), обладают противовирусной активностью (ин терферон, лимфотоксин) и т. д.
Основными методами разграничения и выявления морфологи чески сходных, но функционально разнородных лимфоцитоподобных Т- и В-клеток и их субпопуляций в настоящее время являют ся иммунологические, основанные на определении рецепторов клеточной поверхности. Для идентификации В-лимфоцитов по по верхностным иммуноглобулинам используется метод иммунофлуоресценции. Идентификация Т-лимфоцитов in vitro основана на их способности к спонтанному розеткообразованию с эритроцитами барана (метод бласттрансформации с фитогемаагглютинином или конканавалином А).
Кровяные пластинки
Кровяные пластинки — безъядерные элементы сосудистой крови млекопитающих. Это небольшие цитоплазматические фрагменты,
отделившиеся от мегакариоцитов красного костного |
мозга. |
В 1 мкл крови 250000—350 000 кровяных пластинок. |
В крови |
птиц сходными по функции элементами являются небольшие клет ки с ядром, называемые т р о м б о ц и т а м и .
При световой микроскопии в мазках окрашенной крови обыч но обнаруживают скопления кровяных пластинок. Отдельные пластинки имеют вид дискообразных структур величиной 1—- 3 мкм, со слабобазофильной и гомогенной наружной частью — гиаломер (греч. гиалос — стекло; мерос — часть) и с окрашенны ми азуром зернами — грануломер. При электронной микроскопия в кровяных пластинках различают поверхностную мембрану, по крытую снаружи гликокаликсом, содержащим кислые гликозамипогликаны. Непосредственно под мембраной и параллельно ей располагаются микротрубочки и актиновые микрофиламенты; со кратительные свойства последних проявляются при активация пластинок. Во внутренней зоне находятся несколько митохондрий и два типа гранул: плотные гранулы, в которых содержатся АТФ, катехоламины и серотонии, и так называемые а-гранулы лизосомной природы. Кроме того, в пластинках обнаружены относи тельно крупные по диаметру канальцы, образованные путем впячивания поверхностной мембраны (рис. 100).
В сосудистой крови пластинки существуют около 9—10 суток, после чего происходит их фагоцитоз, главным образом макрофа гами селезенки.
Кровяные пластинки имеют важнейшее значение в обеспече нии основных этапов остановки кровотечения — г е м о с т а з а . На месте повреждения эндотелия стенки сосуда и обнажения базальной мембраны происходит оседание и агрегация пластинок. По следующая их активация сопровождается изменением формы (пластинки становятся шаровидными) и выделенпем ряда соеди нений, содержащихся в гранулах (тромбоцитарные факторы),
234 |
135 |
|
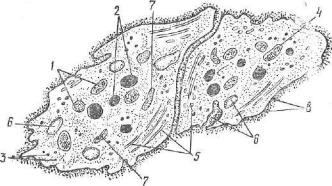
Рис. 100. Схема ультрамикроскопического строения кровяных пластинок:
I — а гранулы; 2 — плотные гранулы; 8 — гликоген; 4 — митохондрии; 5 — микро трубочки; б — трубочки, связанные с поверхностью; 7 — плотные трубочки; 8 — гликокаликс.
которые ускоряют агрегацию пластинок. Кровяные пластинки крови кроликов выделяют значительное количество гистамина. В результате агглютинации все новых и новых пластинок образу ется сгусток (белый тромб), препятствующий выхождению фор менных элементов крови из поврежденного сосуда. Вследствие изменения поверхности кровяных пластинок активируются факто ры свертывания, находящиеся в плазме крови, которые приводят к появлению нерастворимого фибрина, заполняющего простран ства между коагулированными пластинками. В последующей ре тракции кровяного сгустка имеет значение содержащийся в плас тинках т р о м б о с т е н и н (сократительный белок).
ЛИМФА
Лимфа — почти прозрачная желтоватая жидкость, находящаяся в полости лимфатических капилляров и сосудов. Образование ее обусловлено переходом составных частей плазмы крови из крове носных капилляров в тканевую жидкость и поступлением их вме сте с продуктами обмена веществ, выделяемыми клетками соеди нительной ткани в лимфатические капилляры. В образовании лимфы существенное значение имеют взаимоотношение гидроста тического и осмотического давления крови и тканевой жидкости, проницаемость стенки кровеносных капилляров, а также физикохимическое состояние основного вещества соединительной ткани.
Лимфа состоит из жидкой части — лимфоплазмы и формен ных элементов. Л и м ф о п л а з м а отличается от плазмы крови меньшим содержанием белков. В лимфе содержится фибриноген, поэтому она также способна свертываться. Главные форменные
элементы лимфы — л и м ф о ц и т ы ; встречаются |
и другие виды |
лейкоцитов. Состав лимфы в различных сосудах |
лимфатической |
системы неодинаков. Различают периферическую лимфу (лимфа лимфатических капилляров и сосудов до лимфатических узлов), промежуточную (лимфа сосудов после прохождения через лимфа тические узлы) и центральную (лимфа грудного и правого лимфа тического протоков), наиболее богатая клеточными элементами.
КРОВЕТВОРЕНИЕ
Постэмбриональное кроветворение (физиологическая регенерация крови)
Кроветворение (гемоцитопоэз) — многостадийный процесс после довательных клеточных превращений, приводящий к образованию зрелых клеток периферической сосудистой крови. В постэмбрио нальный период у животных развитие клеток крови осуществля ется в двух специализированных интенсивно обновляющихся тка нях, относящихся к разновидностям тканей внутренней среды и условно названных миелоидной и лимфоидной. В них постоянно совершается сбалансированный процесс новообразования и гибе ли клеточных элементов. Представлены они многочисленными гемопоэтическими клетками разного типа в комплексе с ретику лярными или эпителиальными элементами и макрофагами.
Вмиелоидной ткани (греч, миелос — мозг) красного костного мозга происходит развитие стволовых кроветворных клеток и всех форменных элементов крови — эритроцитов, гранулоцитов, лим фоцитов, кровяных пластинок, моноцитов.
Влимфоидной ткани, находящейся в тимусе, селезенке, лим фатических узлах, слизистых оболочках внутренних полостных органов, образуются лимфоциты, а также клетки, являющиеся конечными стадиями дифференциации стимулированных Т- и В- лимфоцитов.
С помощью клональных, иммунологических, электронно-мик роскопических, генетических и радиобиологических методов за последние 25 лет получены важные данные, характеризующие ки нетику клеточных популяций в процессе кроветворения. Отраже нием этого явилось построение новых схем кроветворения, в кото рых уточнены ранние стадии гемоцитопоэза, когда разделение клеток по морфологическим признакам еще невозможно. В настоя щее время наиболее признанной является схема кроветворения, предложепиая И. Л. Чертковым и А. И. Воробьевым (1981), в соответствии с которой весь гемоцитопоэз разделен на шесть эта пов и соответственно выделено шесть классов кроветворных кле ток (цв. табл. II).
Исходя из представления, сформулированного более 60 лет назад А. А. Максимовым, о происхождении клеток крови из еди ного источника признано, что родоначальным элементом всех клеток крови является полипотентная с т в о л о в а я к л е т к а (колониеобразующая единица в селезенке — КОЕс ), способная к разнообразным превращениям и обладающая свойством само-
136 |
137 |
|
поддержания (пролиферации без видимой дифференциации) сво его численного состава в течение всей жизни организма. Популя ция стволовых клеток в схеме кроветворения считается клетками первого класса. Во взрослом организме наибольшее количество •стволовых клеток находится в красном костном мозге (на 100 000 -клеток костного мозга приходится около 50 стволовых), из кото рого они мигрируют в тимус, селезенку, а у птиц в фабрициеву сумку.
Пролиферируют и развиваются стволовые клетки в том или ином направлении под влиянием близкодействующих индукторов микроокружения, образуемых клетками стромы — различными клетками ретикулярной (в красном костном мозге, селезенке) или ретикуло-эпителиальной основы (в тимусе). Несмотря на то что стволовая клетка кроветворения способна проделывать около 100 митозов, в нормальных физиологических условиях основная масса стволовых клеток митотически инертна. Усиление их митотической активности и восстановление характерного для крове творной системы данного организма количества стволовых клеток происходят при воздействиях, резко снижающих общую массу гемопоэтических элементов (например, после кровопотерь или воздействия лучистой энергии). Светомикроскопическое и элек тронно-микроскопическое исследование наиболее очищенной фрак ции стволовых клеток показало, что они имеют по своей морфо логии сходство с малыми лимфоцитами.
Ближайшей ступенью превращения стволовой клетки в про цессе кроветворения является второй класс — частично детерми нированных клеток-предшественников двух разновидностей — миелопоэза и лимфопоэза. Это популяция п о л у с т в о л о в ы х к л е т о к с более ограниченными способностями к самоподдержанию. На агаровой культуре эти клетки образуют колонии, поэтому они получили название «колоннеобразующие единицы» — КОЕ. Под тверждено существование клетки-предшественницы гранулоцитарного, эритроцитарного, моноцитарного и мегакариоцитарного рядов гемопоэза (КОЕ — ГЭММ). Интенсивность их размножения и превращения в следующий, третий класс — «унипотентные клетки-предшественницы», обладающие еще меньшими способно стями к самоподдержанию, регулируется действием специфиче ских биологически активных веществ — подтипов.
В настоящее время в третий класс поэтинчувствительных кле ток отнесены клетки, способные к дифференцировке в направле нии как двух ростков — клетка-предшественница грануло- и моноцитопоэза (КОЕ—ГМ), клетка гранулоцито- и эритроцитопоэза (КОЕ—ГЭ), клетка мегакариоцито- и эритроцитопоэза (КОЕ— МГЦЭ), так и клетки, дифференцирующиеся лишь в одном на правлении, — клетка-предшественница гранулоцитов (КОЕ—Г), клетка-предшественница моноцитопоэза (КОЕ—М), клетка-пред шественница эозинофилов (КОЕ—Эо), клетка-предшественница базофилов (КОЕ—Б), клетка-предшественница мегакариоцитов (КОЕ—МГЦ). Что касается лимфопоэза, то еще не получено
подтверждения существования общей (для Т- и В-лимфоцитов) клетки-предшественницы, и она в схеме остается гипотетичной. Однако на основании обнаружения соответствующих клеточных антигенных маркеров выявлены клетки-предшественницы отдель но для Т- и В-лимфоцитов.
Перечисленные выше классы стволовых, полустволовых и унипотентных предшественников имеют лимфоцитоподобный вид и морфологическими методами не распознаются. Если за счет ство ловых клеток происходит качественная регуляция кроветворения, то есть снабжение кроветворной системы всеми видами предшест венников, то на стадии поэтинчувствительных и следующих за ней морфологически распознаваемых стадиях большинство клеток находится в состоянии пролиферации. Именно в этом отделе реа
лизуется |
основная |
количественная |
регуляция |
кроветворения, |
то есть |
обеспечение |
необходимого |
количества |
клеток нужного |
типа в ответ на конкретные потребности организма.
Далее следует четвертый класс клеток типа «бластов» (эритробласты, миелобласты, лимфобласты и т. д.). Все они имеют более крупные размеры (20 мкм и более), ядро с нуклеолами и нежносетчатым хроматином, неширокий ободок беззернистой, слабобазофилыгой цитоплазмы. Несмотря на то что каждый «бласт» развивается в направлении лишь одного определенного типа кле ток, морфологически все они трудно различимы.
Пятый и шестой классы морфологически распознаваемых кле ток — это соответственно класс созревающих (миелоцит, нормоцит и др.) и класс зрелых клеток (эритроциты, гранулоциты и др.). На уровне последних двух классов выявлено принципиаль ное различие в поведении клеток миелоидного и лимфоидного рядов. Если в последних стадиях миелоидного кроветворения раз витие идет вплоть до гибели клеток, то в лимфоидном ряду воз можно превращение морфологически зрелых лимфоцитов в власт ные формы. Однако это происходит под влиянием специфических индукторов — антигенов (антигензависимая бласттрансформация). Таким образом, в основном подтверждается выдвинутое А. А. Максимовым (1927) представление о том, что малый лим фоцит крови не является конечной стадией дифференциации кле ток лимфоидного ряда, а сохраняет способность трансформиро ваться в клетки, способные к митотическому делению.
Развитие эритроцитов (эритроцитопоэз) в красном костном мозге протекает по схеме: стволовая клетка (СК) — полустволо вые клетки (КОЕ—ГЭММ, КОЕ—ГЭ, КОЕ—МГЦЭ) — унипотент ные предшественники эритропоэза (БОЕ—Э, КОЕ—Э) — эритробласт — пронормоцит — нормоцит базофильный — нормоцит полихроматофильный — пормоцит оксифильный — ретикулоцит — эритроцит. До стадии эритробласта клеткам несвойственны характерные отличительные морфологические признаки, как по лагают, они имеют лимфоцитоподобный вид. О свойствах этих клеток судят на основании данных, получаемых главным обра зом методом клонирования в полутвердых средах, содержащих
138 |
139 |
|
агар, метилцеллюлозу и др. Показано, что в обычных условиях эритроцитопоэза непосредственный предшественник — эритропоэтинчувствительиая унипотентная клетка (КОЕ—Э) образуется из клеток, формирующих большие колонии — бурсты, состоящие из нескольких тысяч эритроидных клеток, так называемые бурстообразующие единицы (БОЕ—Э). В условиях повышенной потреб ности в эритроцитах эритроцитопоэз может миновать стадии БОЕ—Э и КОЕ—Э (на схеме обозначено пунктирными линиями).
Конечный период эритроцитопоэза (начиная с эритробластов) сопровождается образованием морфологически распознаваемых клеток. При этом происходят характерные морфологические изме нения: уменьшаются размеры всей клетки, отмечают ее уплотне ние, затем исчезает ядро, изменяется окраска цитоплазмы. Э р и т- р о б л а с т ы — клетки размером от 15 до 25 мкм. Ядро, занимаю щее ее большую часть, содержит мелко распыленный хроматин п 1—3 ядрышка. Образующиеся из эритробластов пронормоциты имеют меньшие размеры (12—18 мкм) и более грубую структуру хроматина ядра (рис. 101). Пронормоциты — интенсивно делящие ся клетки. В результате митотического деления образуются клет ки размером 10—12 мкм, с плотным ядром и интенсивно базо-
Рнс. 101. Электронная микрограмма пронормоцита (по Агееву) i — митохондрии; 2 — рибосомы; а — ядрышко.
фильпой цитоплазмой, в которой становится заметной узкая более светлая перинуклеарная з о н а — б а з о ф и л ь н ы е н о р м о ц и т ы .
Базофилия цитоплазмы обусловлена наличием в ней большого количества РНК, свободных рибосом и полисом, с которыми свя зан синтез белкового компонента гемоглобина. Последний накап ливается сначала в перинуклеарной зоне. Железо, входящее в со став гемоглобина, базофильные нормоциты получают от макро фагов, фагоцитирующих гибнущие эритроциты. В результате деления базофильных нормоцитов появляются еще более мелкие клетки, цитоплазма которых из-за накопленного гемоглобина утрачивает выраженную ранее базофилию и окрашивается как
основными, |
так и кислыми красителями — п о л и х р о м а т о- |
ф и л ь н ы е |
н о р м о ц и т ы . Ядра их, как правило, без ядрышек, |
а вследствие радиального расположения грубых глыбок гетерохроматина имеют вид колеса со спицами. Проделав завершающее деление, полихроматофильные нормоциты превращаются в клет
ки, цитоплазма которых вследствие |
обилия в |
ней гемоглобина |
|
проявляет |
выраженную оксифилию |
— о к с и ф и л ь н ы е нор |
|
м о ц и т ы . |
Ядро их постепенно уменьшается, |
пикнотизируется |
|
(уплотняется) и отсоединяется. Отделившиеся ядра нормоцитов фагоцитируются макрофагами костного мозга. Образуются моло дые эритроциты — р е т и к у л о ц и т ы , поступающие в кровяное русло. В них еще некоторое время сохраняются РНК-содержащие структуры в виде сеточки. В процессе развития морфологически распознаваемые клетки эритроцитарного ряда осуществляют 5—6 митозов.
Установлено, что даже в нормальных условиях кроветворения часть эритробластов (до 10%) не завершает свой цикл развития до эритроцитов и с помощью макрофагов костного мозга разруша ется.
Этот процесс, названный неэффективным эритропоэзом, является одним из физиологически обусловленных механизмов регуляции в системе эритропоэза. Наиболее сильным регулято ром эритропоэза является количество кислорода, доставляемого к тканям и органам. Недостаточное снабжение кислородом стимули рует усиленную выработку эритропоэтииа, посредством которого регулируется интенсивность пролиферации костномозговых пред шественников (преимущественно на уровне БОЕ—Э и КОЕ—Э) эритропоэза. Эритропоэтин — гормон гликопротеидной природы. Считают, что он синтезируется в почках.
Развитие гранулоцитов (гранулон,итопоэз). При развитии гранулоцитов из стволовых клеток красного костного мозга вначале также образуются морфологически нераспознаваемые полустволо вые (КОЕ—ГЭММ; КОЕ—ГМ; КОЕ—ГЭ) и унипотентные пред шественники (КОЕ—Б; КОЕ—Эо; КОЕ—Гн), которые через ста дии распознаваемых клеточных форм (миелобласт, промиелоцит, миелоцит, метамиелоцит, палочкоядерный гранулоцит) превраща ются в зрелые сегментоядерные грапулоциты трех разновидно стей — нейтрофилы, эозинофилы и базофилы.
140 |
141 |
|
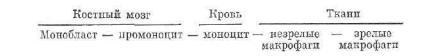
М и е л о б л а с т — первая морфологически идентифицируемая клетка гранулоцитарного ряда —имеет крупное центрально рас» положенное ядро, в котором на фоне диффузно-мелкозернистого хроматина видно несколько ядрышек. Цитоплазма слабо базофильная, в ней можно обнаружить небольшое количество азурофильных гранул. Электронв о-микроскопически в ней выявляются мито хондрии, полирибосомы, элементы гранулярной цитоплазматиче-
ской сети. Разделившись, миелобласт превращается в |
п р о м и е - |
л о ц и т — крупную клетку (20—25 мкм), в которой |
округлое |
ядро часто расположено эксцентрично и содержит 1—2 ядрышка. В элементах пластинчатого комплекса Гольджи промиелоцитов происходит основной процесс формирования азурофильных гра нул. Они в значительном количестве содержатся в цитоплазме и дают положительную реакцию на пероксидазу. После деления из промиелоцита образуются клетки еще меньших размеров и с бо лее плотным ядром, имеющим чаще всего овальную форму, — миелоциты. В цитоплазме миелоцитов наряду с первичной (азурофильной) зернистостью образуются и содержатся вторичные (специфические) гранулы, в соответствии с особенностями кото рых удается отчетливо различать нейтрофильные, эозинофильные
ибазофильные миелоциты.
Вразвитии гранулоцитов миелоциты являются завершающими клетками, способными к митотическому делению, после которого они созревают, последовательно превращаясь в метамиелоциты, палочкоядерные и сегментоядерные клетки. Эти этапы созревания характеризуются некоторыми общими морфологическими измене ниями: уменьшением клетки в размерах, изменением формы ядра от овальной (миелоциты) через бобовидную (метамиелоциты) и палочковидную к расчлененной на дольки (сегментоядерные) и окончательным оформлением в цитоплазме соответствующей спе цифической зернистости. Сегментоядерные гарнулоциты поступа ют из костного мозга в кровяное русло, циркулируют в нем 8— 12 ч, после чего проникают в ткани, где выполняют специфические функции и погибают. Показано, что для эозинофилов тканевая фаза их жизни продолжается около 10 сут, базофилы же погиба ют очень быстро.
Развитие моноцитов (моиоцитопоэз). Клетки моноцитарного ряда образуются в костном мозге из стволовых клеток через ста дии полустволовых клеток (КОЕ—ГЭММ и КОЕ—ГМ), из кото рых возникают унипотентный предшественник (КОЕ—М) и затем монобласты. Немногочисленные м о н о б л а с т ы трудно отличимы от других бластных форм в красном костном мозге. После того как в их цитоплазме сформируются компоненты комплекса Голь джи и образуются азурофильные гранулы, клетки превращаются в промоноциты и моноциты. Последние выходят в кровоток, а затем, проникая в ткани, дают начало незрелым, а позднее зрелым мак рофагам.
Последовательное развитие клеток линии моноцитов — макро фагов из монобластов представлено на схеме.
Костный мозг |
Кровь |
Ткани |
Монобласт — нромоноцит — моноцит — незрелые — зрелые
макрофаги макрофаги Развитие лимфоцитов. Лимфоцитопоэз — один из наиболее
сложных процессов дифференцировки стволовых кроветворных клеток. Важная особенность этого процесса состоит в том, что раз вивается сходная морфологически, но разнородная в функциональ ном отношении клеточная популяция. С участием различных ор ганов поэтапно осуществляется формирование двух тесно связан ных при функционировании линий клеток — Т- и В-лимфоцитов. В красном костном мозге образуются родоначальные лимфоидные клетки, общие как для Т-, так и для В-лимфоцитов. В централь ных лимфоидных органах (тимусе, фабрициевой сумке) лимфоци топоэз зависит от наличия жизнеспособных костномозговых пред шественников. В периферических лимфоидных органах (лимфа тические узлы, селезенка, лимфоидные образования слизистых оболочек) лимфоцитопоэз является антигензависимым процессом.
Развитие кровяных пластинок (тромбошттопоэз). Образование кровяных пластинок происходит в красном костном мозге и свя зано с развитием в нем особых гигантских клеток — м е г а к а р и о ц и т о в . Мегакариоцитопоэз состоит из следующих стадий: •стволовая клетка (СК) — полустволовые клетки (КОЕ—ГЭММ и КОЕ—МГЦЭ) — унипотентные предшественники (КОЕ— МГЦ) — мегакариобласт — промегакариоцит — мегакариоцит. По мере созревания, в результате своеобразной многократной эндометотической репродукции, формируются крупные клетки (40— 50 мкм), содержащие в многолопастном ядре до 32—64 хромосом ных наборов. В развивающихся мегакариоцитах, в цитоплазме образуются система микрофиламентов и микротрубочек, а также специфические гранулы. На заключительных этапах с участием •формирующейся системы из гладких мембран происходит фраг ментация цитоплазмы мегакариоцитов на обособленные участки— кровяные пластинки, которые через стенки синусоидов красного костного мозга попадают в кровоток. После отсоединения пласти нок вокруг оставшегося ядра мегакариоцитов возникает новая цитоплазма. Полагают, что в каждом мегакариоците красного костного мозга совершается циклический процесс развития не скольких поколений кровяных пластинок.
Эмбриональное кроветворение
В течение эмбрионального периода развития организма происхо дит последовательная смена локализации кроветворения. На са мых ранних этапах клетки крови образуются за пределами заро дыша, в мезенхиме стенки желточного мешка, где формируются скопления — к р о в я н ы е о с т р о в к и . Центральные клетки островков округляются, обособляются и преобразуются в стволо вые кроветворные «летки. Периферические клетки островков рас-
142