

gistologia
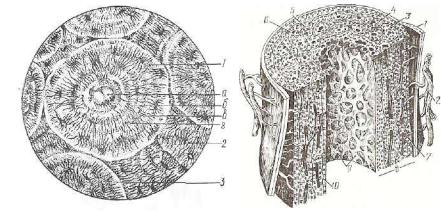
.pdfРис. 113. Ретикулярная ткань в краевом синусе лимфа тического узла:
1 — ретикулярные кпетни; 2 — лимфоциты.
находятся тканевая жидкость и различные свободные клеточные элементы (рис. 113). Ретикулярная ткань образует строму крове творных органов, где в комплексе с макрофагами создает специ фическое микроокружение, обеспечивающее размножение, диффе ренциацию и миграцию разных форменных элементов крови. В небольшом количестве ретикулярная ткань содержится в пече ни и в подэпителиальной соединительной ткани слизистых оболо чек.
Ретикулярные клетки развиваются из мезенхимоцитов и в постэмбриональный период имеют сходство с другими типами механоцитов — фибробластами, хондробластами и др. Они имеют разные размеры и звездчатую форму, обусловленную наличием мно жества отростков. Цитоплазма при окраске препаратов гематокси лином и эозином слабо-розовая. Ядро чаще круглой формы, содер жит 1—2 отчетливо выраженных ядрышка. При электронно-мик роскопическом исследовании выявляют глубокие впячивания ядер ной оболочки. В цитоплазме встречаются свободные полисомы и рибосомы, элементы гладкой эндоплазматической сети, немного численные мелкие митохондрии. Степень развития гранулярной эндоплазматической сети и комплекса Гольджи может быть раз личной. В области контакта отростков соседних клеток находятся десмосомы. Гистохимически для ретикулярных клеток характер на низкая активность эстеразы и кислой фосфатазы и высокая активность щелочной фосфатазы. Ретикулярные клетки практиче ски не делятся и отличаются высокой устойчивостью к воздей ствию ионизирующего излучения.
164
Рис. 114. Схема взаимоотно шения ретикулярной клетки и ретикулярных волокон:
1 — ядро ретикулярной клет ки; 2 — отростки ретикуляр ной клетки; 3 — ретикуляр- _ ные волокна; 4 — эндоплаз- 5 матическая сеть; 5 — мито хондрии.
Ретикулярные волокна — производные ретикулярных клеток и представляют тонкие ветвящиеся волокна, образующие сеть. При окрашивании срезов гематоксилин-эозином ретикулярные волок на не выявляются. Для их обнаружения используют различные варианты импрегнации солями серебра. При электронной микро скопии в составе ретикулярных волокон обнаружены различные по диаметру фибриллы, заключенные в гомогенное плотное меж фибриллярное вещество. Фибриллы состоят из коллагена III типа и имеют свойственную коллагеновым фибриллам поперечную исчерченность — чередование по длине фибриллы темных и свет лых дисков. Периферическое расположение межфибриллярного компонента, содержащего значительное количество полисахаридов (до 4%), обусловливает высокую устойчивость ретикулярных во локон к действию кислот и щелочей и способность восстанавли вать серебро при окраске волокон.
Обнаружены тесные взаимоотношения ретикулярных волокон с ретикулярными клетками — волокна расположены, как правило, в глубоких инвагинациях (впячиваииях) плазмолеммы клеток (рис. 114).
ХРЯЩЕВАЯ ТКАНЬ
Хрящевая ткань — специализированный вид соединительной тка ни, выполняющий опорную функцию. В эмбриогенезе она разви вается из мезенхимы и формирует скелет зародыша, который в последующем в большей части замещается костью. Хрящевая ткань, за исключением суставных поверхностей, покрыта плотной соединительной тканью — надхрящницей, содержащей сосуды, питающие хрящ и его камбиальные клетки.
Хрящ состоит из клеток — хондроцитов и межклеточного веще ства. В соответствии с характеристикой хондроцитов и особенно межклеточного вещества различают три вида хрящей: гиалиновый, эластический и волокнистый.
Г и с т о г е н е з х р я щ е в о й т к а н и . В процессе эмбрио нального развития зародыша мезенхима, интенсивно развиваясь, образует островки плотно прилегающих друг к другу клеток протохондральной (предхрящевой) ткани (рис. 115). Ее клетки ха рактеризуются высокими значениями ядерно-цитоплазматических отношений, мелкими, плотными митохондриями, обилием свобод-
165
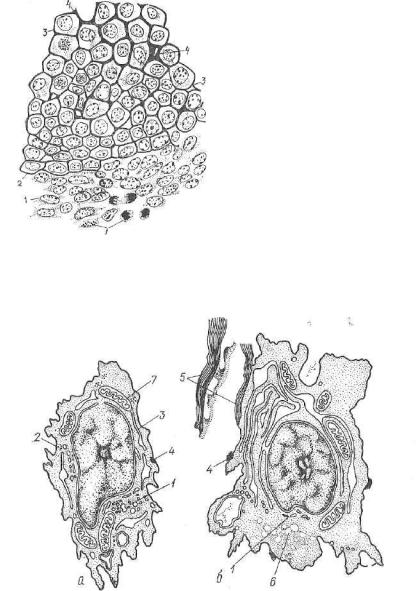
Рис. 115. Развитие гиалинового хря ща из мезенхимы:
1 — мезенхима; 2 — ранняя стадия раз вития хряща; 3 — более поздняя стадия дифференцировки хряща; 4 — промежу точное вещество развивающегося хряща.
ных рибосом и слабым развити ем гранулярной эндоплазматической сети. Комплекс Гольджи в клетках протохондральной тка ни рассредоточен в виде неболь ших цистерн и пузырьков (рис. 116). По мере дифференцировки хондробластов они включаются в процессы синтеза макромолекулярных соединений межкле точного вещества развивающе
гося хряща, соответственно изменяется их синтетический и сек реторный аппарат. Увеличивается объем цитоплазмы и соот ветственно уменьшается показатель ядерно-цитоплазматичес- ких отношений. Возрастает количество цистерн гранулярной зндоплазматической сети. Комплекс Гольджи сосредоточивает-
Рис. 116. Схема последовательных изменении ультраструктурнои организа ции клеток (а, б) при гистогенезе хрящевой ткани млекопитающих (по Кодман, Портер):
1 — комплекс Гольджи; г — свободные рибосомы; 8 — эндоплазматическая сеть гранулярная; 4 — уплотненные участки цитоплазмы в области выведения макро молекул; 5 — коллагеновые фибриллы; 6 — область концентрации гликогена; 7 — -митохондрии.
ся вокруг ядра и расширяются его размеры. Увеличивается объем митохондрий преимущественно за счет возрастания мас сы их матрикса. Наблюдается выведение в окружающее межкле точное вещество содержимого вакуолей клеток. В межклеточное вещество секретируют тропоколлаген и неколлагеновые белки, а затем гликозаминогликаны и протеогликаны. Формируется пер вичная хрящевая (прехондральная) ткань.
Дифференцированный хондробласт морфологически характе ризуется хорошо развитой гранулярной эндоплазматической сетью и обширным комплексом Гольджи. В цитоплазме клеток много включений гликогена. Увеличение массы хрящевого зачат ка в процессе эмбриогенеза идет как за счет увеличения количе ства межклеточного вещества, так и за счет размножения хондро бластов.
По ме'ре накопления |
межклеточного вещества клетки разви |
||
вающегося хряща изолируются в |
отдельных |
полостях (лакунах) |
|
и дифференцируются в |
зрелые |
хрящевые |
клетки — х о и д р о- |
ц и ты. |
|
|
|
Дальнейший рост хрящевой ткани обеспечивается продолжаю щимся делением хондроцитов и формированием между дочерними клетками межклеточного вещества. В более поздних стадиях раз вития ткани образование межклеточного вещества замедляется. Дочерние клетки, оставаясь в одной лакуне или отделяясь друг от друга лишь топкими перегородками основного вещества, обра зуют характерные для зрелого хряща изогенные группы клеток (от isos — равный, одинаковый, genesis — происхождение). В даль нейшем рост хрящевой ткани обеспечивается как увеличением ее массы размножением клеток хрящевой закладки и соответст венно формированием межклеточного вещества — ее интерстициальным ростом, так и продолжающимся развитием хряща за счет внутреннего — камбиального слоя надхрящницы, клетки которой,, размножаясь и дифференцируясь в хондроциты, обусловливают аппозиционный рост ткани.
По мере дифференцировки хрящевой ткани интенсивность раз множения клоток падает, ядра пикгютизируются, ядрышковый аппарат родуцируется.
Г м а л и и о п ы ii x p я щ. Во взрослом организме гиалиновый хрящ входит в состав ребер, грудины, покрывает суставные по верхности костей, образует хрящевой скелет воздухоносных пу тей: носа, гортани, трахеи, бронхов.
Клетки хрящевой ткани. Клетки хряща — хондроциты — раз личных его зон характеризуются специфическими особенностями формы, положения и высоты дифференцировки. Так, непосредст венно под надхрящницей локализованы незрелые хрящевые клет ки — х о н д р об л а с т ы . Они овальной формы и ориентированы длинной осью параллельно поверхности хряща. Цитоплазма этих клеток богата рибонуклеиновой кислотой, что определяет ее базофилию. В более глубоких зонах хряща хондриоциты округляются или имеют неправильную многоугольную форму, их объем увели-
167
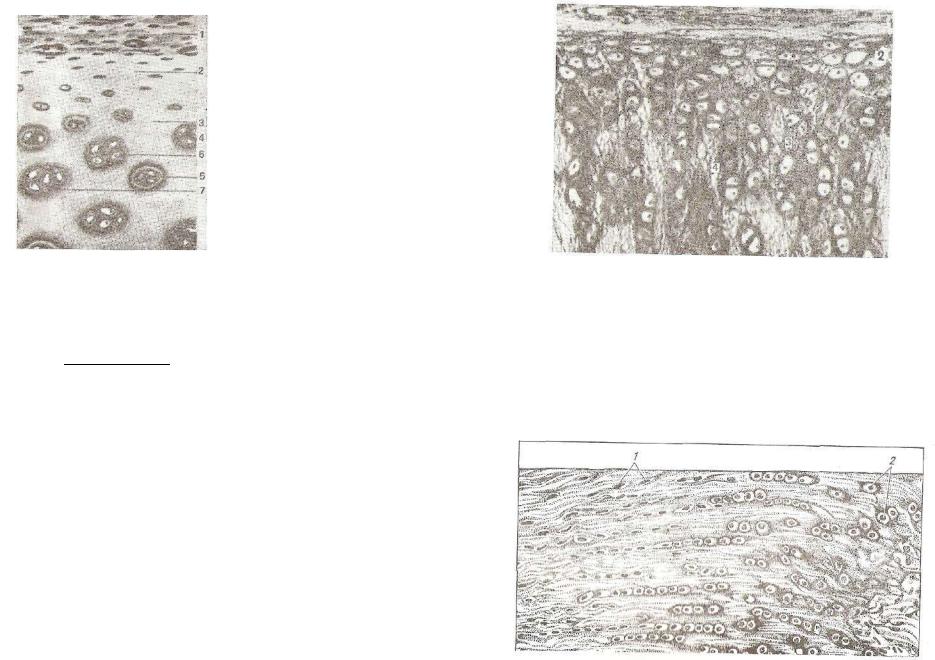
Ряс. 117. Гиалиновый хрящ:
1 — надхрящница; 2 — зона хряща с молодыми хрящевыми клетками; 3 — ' основное вещество; 4 — высокодифференцированные хрящевые клетки; 5 — капсула хрящевых клеток; е — изогенные группы хрящевых клеток; 7 — базофильное основное вещество вокруг хрящевых клеток.
ЧЙарг |
ч |
£\^ \ _ |
' |
^fc-tj |
чивается. В цитоплазме интен- |
|
•jftgflllj»; |
^^^^§^^S^~^7:° |
|
сивно развивается гранулярная |
|||
;*•*•**" »^ |
* й$-й"|~-~- ~% |
эндоплазматическая сеть. Ко мп- |
||||
л^1 *- |
•—~—»--JWit-^---»-^a |
леке Гольджи |
увеличивается в |
|||
|
|
;\puZf |
' 'jiblp'o |
размерах. В митохондриях воз- |
||
р: , |
|
£. >f%i |
* |
* | 1 | < |
растает объем матрикса, в цито- |
|
i jd |
^f******* |
г |
, |
плазме клеток |
накапливаются |
|
f*.w**_ |
|
|
|
- Л |
включения гликогена и липои |
|
дов. Клетки здесь располагаются группами в одной или смежных лакунах, образуя характерные для хряща «изогевные группы» клеток, то есть группы, образовав шиеся повторным делением одного хондроцита (рис. 117—4).
Центрально расположенные з р е л ы е х о н д р о ц и т ы имеют крупные округлые ядра с четко выраженным ядрышком. Глыбки хроматина в них преимущественно сосредоточены на внутренней поверхности ядерной оболочки. Цитоплазма зрелых хрящевых клеток богата органеллами. Клеточный центр локализован около ядра. Комплекс Гольджи, агранулярная и гранулярная эндоплаз матическая сеть хорошо развиты, что свидетельствует об активно сти процессов синтеза компонентов межклеточного вещества: его белков, гликозамиигликанов, протеогликанов. Цитоплазма клеток содержит включения гликогена и липидов.
Межклеточное вещество гиалинового хряща содержит до 70% сухого веса фибриллярного белка коллагена и до 30% аморфного вещества, в состав которого входят сульфатированные и несульфатированные гликозамингликаны, протеогликаны, липиды и неколлагеновые белки. В отличие от коллагеновых волокон дру гих видов соединительной ткани коллагеновые фибриллы хряща тонки и не превышают 10 нм в диаметре.
Ориентация волокон межклеточного вещества определяется характерными для каждого хряща закономерностями механиче ской напряженности. В периферической зоне хряща они ориенти рованы параллельно поверхности, тогда как в глубокой зоне его
их положение меняется в зависимости от специфичности механи
168
ческих нагрузок. Гликозамингликаны, гликопротеиды и неколла-
геновые белки закономерно распределяются в межклеточном ве ществе,ночныеоксифкрасильнтекчтеткоями. Концентрация.обусловливаеверетенообразноВ периферичесглиспецифкой замйформы,зонеиигликановчностьхряща,межклеточноег вышсодержащвзаимодзонвеществйствия«клеодио-
Рис. 118. Эластический хрящ ушной раковины:
I — надхрящница; 2 — молодые хрящевые клетки: з группы хрящевых клеток; 4 — эластические волокна.
точных территорий», вокруг изогенных групп клеток центральной области хряща, о чем свидетельствует их базофилия.
Обмен веществ хряща обеспечивается циркуляцией тканевой жидкости межклеточного вещества, которая составляет до 75% оощей массы ткани. Макромолекулы гликозамиигликанов и про теогликанов, удерживая большое количество воды, определяют его механические свойства.
Рис. 119. Волокнистый хрящ на месте прикрепления сухожилия к боль шой берцовой кости:
1 — сухожильные клетки; г — хрящевые клетки.
169
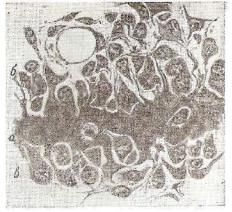
Э л а с т и ч е с к и й х р я щ образует скелет наружного уха, слухового прохода, евстахиевых труб, клиновидных и рожковых хрящей гортани. В отличие от гиалинового хряща в состав его межклеточного вещества, кроме аморфного вещества и коллагеновых фибрилл, входнт плотная сеть эластических волокон, которая на периферии переходит в ткань надхрящницы. Клетки его иден тичны клеткам гиалинового хряща. Они также образуют группы
и лишь под надхрящницей лежат одиночно |
(рис. 118). |
В о л о к н и с т ы й х р я щ локализуется |
в составе межпозво |
ночных дисков, круглой связки бедра, в симфизах лобковых костей, в области прикрепления сухожилия к костям. Межклеточное ве щество волокнистого хряща содержит грубые пучки параллельно ориентированных коллагеновых волокон. Клетки хряща образуют изогенные группы, вытянутые в обособленные цепочки между пучками коллагеновых волокон (рис. 119). Этот вид хряща в основном представляет собой переходную форму между гиалино вым хрящом и плотной соединительной тканью. Регенерация хряща обеспечивается надхрящницей, клетки которой сохраняют камбиальность.
КОСТНАЯ ТКАНЬ
Костная ткань, как и другие виды соединительной ткани, разви вается из мезенхимы, состоит из клеток и межклеточного веще ства, выполняет функцию опоры, защиты и активно участвует в обмене веществ организма. Кости скелета, черепа, грудной клетки, позвоночников обеспечивают механическую защиту органов цен тральной нервной системы и грудной полости. В губчатом вещест ве костей скелета локализован красный костный мозг, здесь осу ществляются процессы кроветворения и дифференцировки клеток иммунной защиты организма. Кость депонирует соли кальция, фосфора и др. В совокупности минеральные вещества составляют 65—70% сухой массы ткани, преимущественно в виде его фосфор ных и углекислых соединений (солей). Кость активно участвует в обмене веществ организма, что определяет ее способность зако номерно перестраиваться, отвечая на изменяющиеся условия его жизнедеятельности, динамику обмена веществ в связи с возрастом, диетой, активностью функции желез внутренней секреции и др.
Клетки костной ткани. Костная ткань содержит четыре раз личных вида клеток: остеогенные клетки, остеобласты, остеоциты
и остеокласты. |
|
О с т е о г е н н ы е к л е т к и — клетки |
ранней стадии специ |
фической дифференцировки мезенхимы в |
процессе остеогенеза. |
Они сохраняют потенцию к митотическому делению. Характеризу ются овальным, бедным хроматином ядром. Их цитоплазма слабо окрашивается основными или кислыми красителями. Локализуют ся эти клетки на поверхности костной ткани: в надкостнице, эндоосте, в гаверсовых каналах и других зонах формирования костной ткани. Остеогенные клетки размножаются и, дифференцируясь,
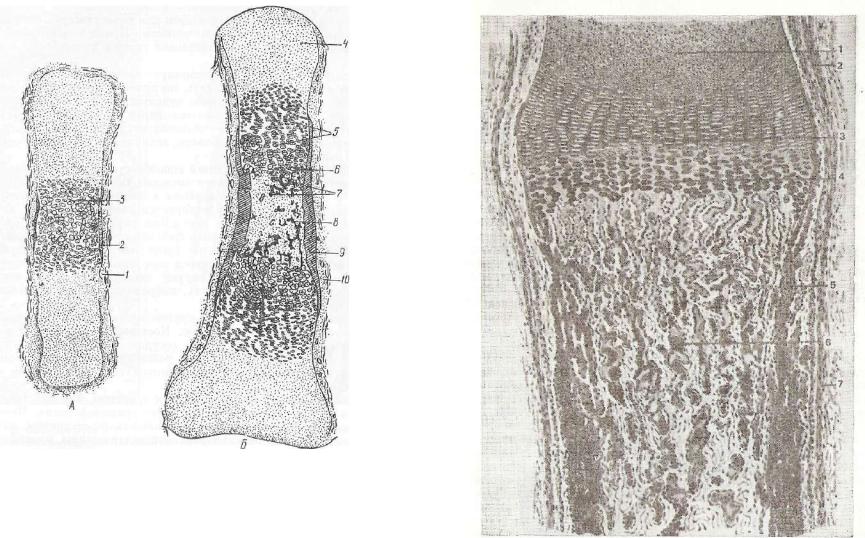
Рис. 120. Развитие кости в ме зенхиме (по Петерсену):
о |
— новообразующееся |
межкле |
точное вещество костной ткани; |
||
б |
— остеобласты. |
|
бластов, обеспечивающих °^^^^^^Ш^^Щ «. -< рост и перестройку костно- ;^^^^Щ^^^Е^^^^^^^^^^^^^^р/:'
го скелета. |
|
|
- ^ ^ ^ Э Д w |
«^' Щ |
f S/ |
Остеобласты |
— |
^ M s A ^ l ^ ^ М^^^§$!%Ш Щ*я |
|||
клетки, продуцирующие |
^ * ;*? /^ |
'~ Г |
^ •* , |
||
органические |
элементы |
- З ^ б р ^ Т З Й ж " ' |
|||
межклеточного |
|
вещества |
^етйЁм^Ш^да^^*^ ^ж^^Мчл^Л |
||
костной ткани: |
коллаген, |
|
|
|
|
гликозамингликаны, белки и др. Это крупные клетки кубической или призматической формы, расположенные по поверхности форми рующихся костных балок. Их тонкие отростки анастомозируют друг с другом. Ядра остеобластов округлые с крупным ядрышком,, расположены эксцентрично. Цитоплазма содержит хорошо разви тую зернистую эндоплазматическую сеть и свободные рибосомы, что определяет ее базофилию (рис. 120, 121, 122). Комплекс Гольджи рассредоточен в цитоплазме клеток между ядром и развиваю щейся костью. Многочисленные митохондрии овальной формы. Для цитоплазмы остеобластов специфична положительная реак ция на активность щелочной фосфатазы.
О с т е о ц и т ы — клетки костной ткани — лежат в особых по лостях межклеточного вещества — л а к у н а х , соединенных меж ду собой многочисленными к о с т н ы м и к а н а л ь ц а м и . Остео циты имеют соответствующую лакуне форму уплощенного овала (22—55 мкм длины и 6—15 мкм ширины). Их многочисленны© тонкие отростки, распространяясь по костным канальцам, анасто мозируют с отростками соседних клеток. Система лакун и кост ных канальцев содержит тканевую жидкость и обеспечивает уро вень обмена веществ, неосбходимый для жизнедеятельности кост ных клеток (рис. 123, 124). Морфологическая организация цито плазмы остеоцитов соответствует степени их дифференцировки. Молодые формирующиеся клетки по составу органелл и степени их развития близки к остеобластам. В более зрелой кости цито плазма 'клеток беднее органеллами, что свидетельствует о сни жении уровня обмена веществ, в частности синтеза белков.
О с т е о к л а с т ы — крупные, многоядерные клетки, от 20 до* 100 мкм в диаметре. Остеокласты находятся на поверхности кост ной ткани в местах ее резорбции. Клетки поляризованные. Поверх ность их, обращенная к резорбируемой кости, имеет большее количество тонких, плотно расположенных, ветвящихся отрост
ков, |
образующих |
в совокупности |
г о ф р и р о в а н н у ю к а е м к у |
(рис. |
125). Здесь |
секретируются и |
сосредоточиваются гидролити- |
170
171
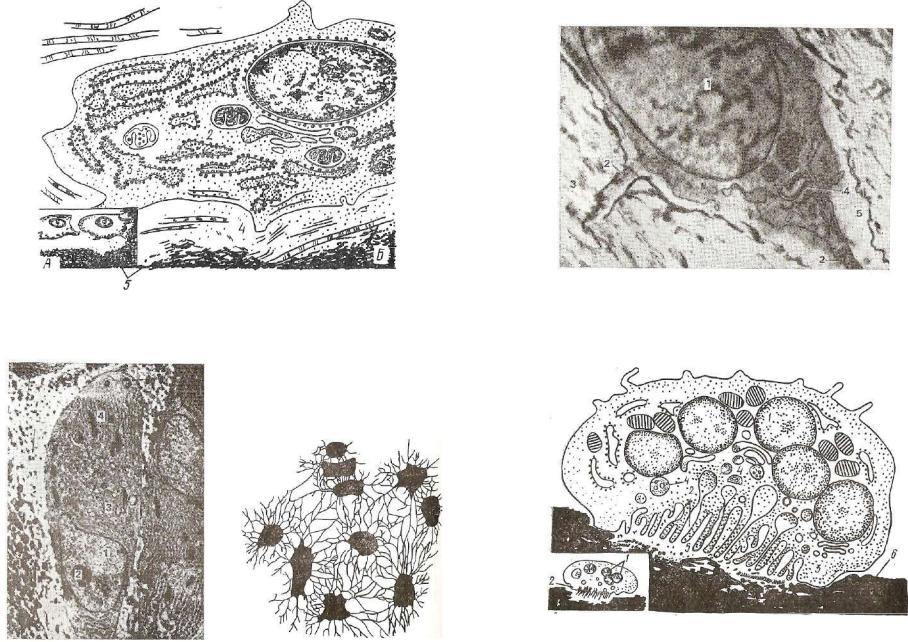
|
|
Рис. 124. Электронная микрофотограмма остеоцита |
(ув. 16 000): |
Рис. 121. Схема строения остеобласта: |
|
1 •— ядро; 2 — отростки остеоцита; 3 — основное обызвествленное веще |
|
1 — ядро; г — ци |
ство, окружающее остеоцит; 4 — альфа-цитомембраны |
эргастоплазмы; |
|
А — на светооптическом; Б — на субмикроскопическом уровне; |
о — основное необызвествленное вещество, непосредственно прилежащее |
||
топлазма; з — развитие гранулярной эндоплазматическои сети; |
4 — остеоид; 5 — |
к остеоциту (по Даллей и Спиро), |
|
минерализованное вещество костной ткани. |
|
|
|
Рис. 122. Электронная микрофотоРис. 123. Костная пластинка из реграмма остеобласта: шетчатой кости белой мыши: видны 1 — ядро; 2 — ядрышко; 8 — цитоплазклетки и межклеточное вещество. матическая сеть; 4 — митохондрии.
ческие ферменты, участвующие в процессах разрушения кости. Область гофрированной каемки граничит с окружающей ее зоной поверхности клетки, плотно прилегающей к резорбцируемой кости с в е т л о й з о н о й , почти не содержащей органелл. Цитоплазма центральной части клетки и ее противоположного полюса содер жит многочисленные ядра (до 100 ядер), несколько групп струк тур комплекса Гольджи, митохондрии, лизосомы. Ферменты лизосом, поступающие в зону гофрированной каемки, активно участ вуют в резорбции кости. Гормоны паращитовидной железы (ПТГ), усиливая процессы секреции ферментов лизосом, стиму лируют резорбцию кости. Кальцитонин щитовидной железы сни жает активность остеокластов. Отростки гофрированной каемки в этих условиях сглаживаются, и клетка отделяется от поверхности кости. Резорбция кости замедляется.
Межклеточное вещество костной ткани состоит из коллагеновых волокон и аморфного вещества: гликопротеидов, сульфатированных гликозамингликанов, белков и неорганических соедине ний — фосфата кальция, гидроапатита и различных микроэлемен тов (медь, цинк, барий, магний и др.)- 97% всего кальция организма сосредоточено в костной ткани. В соответствии со струк турной организацией межклеточного вещества различают грубоволокнистую кость и пластинчатую.
Грубоволокнпетая кость характеризуется значительным диа метром пучков коллагеновых фибрилл и разнообразием их ориен тации. Она типична для костей ранней стадии онтогенеза живот ных и некоторых участков скелета взрослых: зубных альвеол,, костей черепа вблизи костных швов, костного лабиринта внутрен него уха, области прикрепления сухожилий и связок. В пластин чатой кости коллагеновые фибриллы межклеточного вещества не образуют пучков. Располагаясь параллельно, они формируют слои — костные пластинки толщиной 3—7 мкм. Смежные пластин ки всегда имеют различную ориентацию фибрилл. В пластинках закономерно расположены клеточные полости — л а к у н ы и со единяющие их к о с т н ы е к а н а л ь ц а , в которых лежат костные клетки — остеоциты и их отростки (рис. 126). По системе лакун и костных канальцев циркулирует тканевая жидкоеть, обеспечи вающая обмен веществ в ткани.
В зависимости от положения костных пластинок различают губчатую и компактную костную ткань. В г у б ч а т о м в е щ е с т ве, в частности в эпифизах трубчатых костей, группы костных пластинок располагаются под разными углами друг к другу в со ответствии с направлением основных механических нагрузок дан ного участка скелета. Ячеи губчатого вещества кости содержат красный костный мозг. Оно обильно снабжается кровью и актив но участвует в минеральном обмене организма.
В к о м п а к т н о м в е щ е с т в е группы костных пластинок 4—15 мкм толщиной плотно прилежат друг к другу. В соответ ствии с особенностями васкуляризации и локализации камбиаль ных клеток кости — остеобластов в компактном веществе диафизов
Рис. 126. Система остеопов пластин чатой костной ткани (гистологичес кий препарат декальцинированной трубчатой кости. Поперечный срез):
1 — остеон; о — канал остеона с кро веносными сосудами; б — костные пла стинки; в — костные лакуны (полости); г — костные канальцы; 2 — система вставочных пластинок; з — резорбционная (спайная) линия.
трубчатых костей формируется три слоя: наружная общая систе ма пластинок, остеонный слой, содержащий остеоны и вставоч ные системы костных пластинок, и внутренняя общая (окружаю щая) система. Пластинки наружной общей системы формируются остеобластами надкостницы, при этом часть остеобластов превра щается в остеоциты и включается во вновь образованную костную ткань. Костные пластинки наружной общей системы следуют параллельно поверхности кости. Через этот слой кости из надкост ницы проходят п р о б о д а ю щ и е к а н а л ь ц а , несущие в кость кровеносные сосуды и грубые пучки коллагеновых волокон, заму рованные в нее при формировании наружных общих пластинок (рис.127).
В остеонном слое трубчатой кости каналы остеона, содержащие кровеносные сосуды, нервы и сопровождающие их соединитель нотканные элементы, анастомозируя друг с другом, в основном ориентированы продольно. Системы трубкообразных костных плас тинок, окружающие эти каналы, — о с т е о н ы содержат от 4 до 20 пластинок. На поперечных срезах компактного вещества труб чатых костей они определяются как чередующиеся более светлые волокнистые (с циркулярным положением волокон) и более тем ные зернистые слои в соответствии с ориентацией коллагеновых фибрилл межклеточного вещества. Остеоны отграничены друг от друга ц е м е н т н о й л и н и е й основного вещества. Между остеопами включены вставочные, или промежуточные, системы кост ных пластинок, представляющих собой части ранее сформирован
174 |
ие |
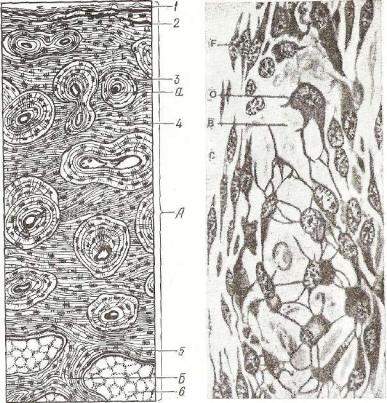
Рис. 128. Пластинчатая кость:
А — плотное (компактное) вещество ко сти; 1 — надкостница; 2 — наружные общие пластинки; 3 — остеоны; а — канал остеона; 4 — система вставочных пластинок; 5 — внутренние общие пла стинки; Б — губчатое вещество кости; 6 — желтый костный мозг.
ных остеонов, сохранившиеся в процессе перестройки кости. По следние очень разнообразны по размерам, форме и ориентации
(рис. 128).
Внутренняя общая (окружающая) система костных пластинок граничит с эндоостом костной полости и представлена пластинка ми, ориентированными параллельно поверхности костно-мозгово-
го канала.
Г и с т о г е н е з к о с т и . Кость, как и другие виды соединитель ной ткани, развивается из мезенхимы. Различают два вида остеогоиоэа: ионосродстношю из мезенхимы и путем замещения костью
имбрИОИПЛЫЮГО чрии1,;\.
176
Развитие кости из мезенхимы — интермембранозиое окостене ние. Этот вид остеогенеза характерен для развития грубоволокнистой кости черепа п нижней челюсти. Процесс начинается с интенсивного развития соединительной ткани и кровеносных сосу дов.
Мезенхимные клетки, анастомозируя между собой отростка ми, в совокупности образуют сеть, погруженную в аморфное меж клеточное вещество, содержащее отдельные пучки коллагеновых волокон. Клетки, оттесненные межклеточным веществом на по верхность такого остеогенного островка, становятся базофильными и дифференцируются в остеобласты, активно участвующие в остеогенезе (рис. 129).
Отдельные клетки, утрачивая способность синтеза межклеточ ного вещества, при активности смежных остеобластов замуровы ваются в него и дифференцируются в остеоциты. Межклеточное вещество молодой кости импрегнируется фосфатом кальция, кото рый накапливается в кости вследствие распада глицерофосфата крови под действием выделяемой фибробластами щелочной фосфатазы. Освобождающийся остаток фосфорной кислоты реагирует с хлоридом кальция. Образующиеся при этом фосфат кальция и углекислый кальций импрегнируют основное вещество кости. Окружая формирующуюся кость, эмбриональная соединительная ткань образует периост.
В последующем первичная грубоволокнистая костная ткань замещается пластинчатой костью. Костные пластинки при этом образуются вокруг кровеносных сосудов, формируя первичные остеоны. Со стороны надкостницы развиваются наружные общие системы костных пластинок, ориентированные параллельно' по верхности кости.
Энхондральное окостенение. Кости туловища, конечностей, ос нования черепа формируются на месте хрящевой ткани. Началопроцесса характеризуется перихондральным окостенением, начи нающимся усиленной васкуляризацией надхрящницы, пролифера цией и дифференциацией ее клеток и межклеточного вещества, в- том числе остеобластов.
В трубчатых костях этот процесс начинается в области диафиза формированием под надхрящницей сети перекладин грубоволокнистой кости — костной манжетки (рис. 130). По мере разви тия периостальной кости в середине ее хрящевой модели в цеп> т р е о к о с т е н е н и я хрящевая ткань закономерно изменяется. К*летки хряща прогрессивно увеличиваются в размерах, обогаща ются гликогеном и васкуляризуются. Их ядра сморщиваются. Кле точные полости увеличиваются. В области диафиза формируется иона пузырчатого хряща (рис. 131). Соединительная ткань над костницы, проникая между перекладинами костной манжетки,, шюсит в зону дегенерирующего хряща различно дифференцироишшые мезенхимные клетки как гемопоэтического ряда, так и дифференцирующиеся клетки костной ткани: остеокласты и остео бласты.
1!2 Зпказ W» 908 |
17? |
В смежных зонах хрящевого зачатка кости клетки, размно жаясь, образуют расположенные параллельными рядами, продоль но ориентированные «клеточные колонки». Клетки в колонке раз граничены тонкими перегородками основного вещества. Межкле точное вещество между колонками клеток, уплотняясь и кальцефицируясь, формирует «хрящевые балки». Эндохондральное •окостенение распространяется от диафиза хрящевой закладки к его эпифизам, соответственно в составе клеточных колонок можно
178
Рис. 131. Эпхондральное и перихондральное развитие кости:
1 — остеобластический слой надкостницы; 2 — фиброзный слой надкостницы; S — перихондриальная костная манжетка; 4 — клеточные колонки; 3 — остеоцитыи в — остеобласты; 7 — остекласт.
выделить наиболее удаленную от диафпза зону пролиферации^ клеток (за которой ближе к диафизу следуют зоны созревания их)г гипертрофии, дистрофии и распада. В образующиеся при этом' лакуны врастают кровеносные сосуды с остеогенными клетками. По мере дифференцировки остеобластов они локализуются на-
12* |
179* |
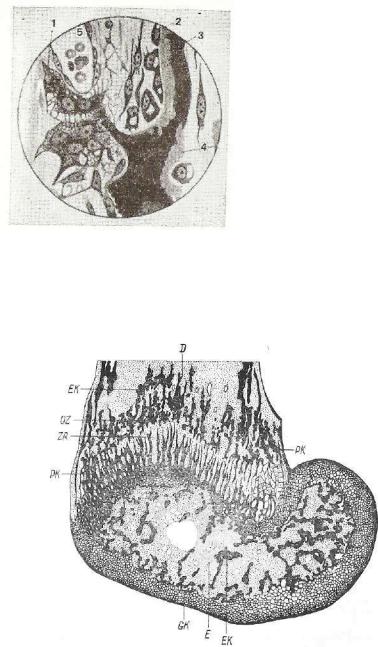
Рис. 132. Энхондральное развитие кости:
1 — остеокласт; г — остеобласт; 3— остатки обызвествленного х^яща; 4— новообразованная кость; 5 — крове носный сосуд.
стенках лакун и, продуцируя
Iмежклеточное вещество ко сти, формируют на поверхно сти сохранившихся хрящевых пластинок костную ткань. Процесс замещения хряща костной тканью называется
*& энхондральным |
окостенени |
ем (рис. 132). |
|
Одновременно с развитием энхондральной кости |
со стороны |
надкостницы идет активный процесс перихордального остеогенеза, формирующий плотный слой периостальной кости, распространя ющейся по всей ее длине до эпифизарной пластинки роста. Периостальная кость представляет компактное вещество кости скелета. В отличие от грубоволокнистой кости манжетки у нее строение
Рис. 133. Срез через эпифиз бедренной кости 4-недельной мыши v(по Ша феру):
? ш ~ . д ^ и з ' £ — эпифиз; |
ЕК — энхондральная кость эпифиза; GK — суставной |
|||
^3L^%bZPSS&5S&£K |
- |
перихондриальная |
кость |
JSASi |
180
типичной пластинчатой кости.с характерными системами костных: пластинок, выраженными в разной степени в зависимости от вида животного и специфичности отдельных костей скелета.
Позднее центры окостенения появляются в эпифизах кости. Развивающаяся здесь костная ткань замещает хрящевую ткань всего эпифиза. Последняя сохраняется лишь на суставной поверх ности ив э п и ф и з а р н о й п л а с т и н к е р о с т а , отграничи вающей эпифиз от диафиза (рис. 133) в течение всего периода рос та организма до половой зрелости животного.
Надкостница (периост) состоит из двух слоев. Ее внутренний слой содержит коллагеновые и эластические волокна, остеоблас ты, остеокласты и кровеносные сосуды. Последние проникают че рез питательные отверстия кости в костную ткань и в костный мозг. Наружный слой надкостницы образован плотной соедини тельной тканью. Она непосредственно связана с сухожилиями мышц и коллагеновыми волокнами связок. Отдельные пучки коллагеновых волокон надкостницы непосредственно включены в костную ткань в виде «прободающих» волокон, обеспечивающих механическую прочность связи надкостницы с костью.
Эндоост — слой соединительной ткани, выстилающий костно мозговой канал. Он содержит остеобласты и тонкие пучки коллагеновых волокон, переходящих в ткань костного мозга.
МЫШЕЧНЫЕ ТКАНИ
Ткани, характерным свойством которых является способность к произвольному и непроизвольному односторонне направленному сокращению, относят к мышечным. Их сокращение, сопряженное с затратой большого количества энергии, образующейся при гид ролизе АТФ, осуществляется с участием особого сократительного аппарата.
В филогенезе развитие этих тканей протекало в связи с не обходимостью активного перемещения многоклеточных организ мов с целью отыскания более благоприятной среды для сущест вования. Фило-онтогенетическое становление мышечных тканей протекало в тесной связи с нервной тканью. Структурное выра жение этой связи проявляется в наличии плазматических мем бран, сходных по своим морфофуикциональным свойствам. Они участвуют в образовании волн деполяризации, лежащих в основе возбуждения и сокращения мышц.
Сократительный аппарат у мышечных тканей белковой при роды, представлен системой тонких нитей (филаментов), распо ложенных по длинной оси их основных структур. Структурами сократительных тканей могут быть мышечные клетки и мышеч ные волокна, что обусловлено местом их нахождения и функ цией.
Единого источника эмбрионального развития у этих тканей нет. ИМИ являются мезенхима, миотомы сегментированной части мезодермы, висцеральный листок спланхнотома и др.
181
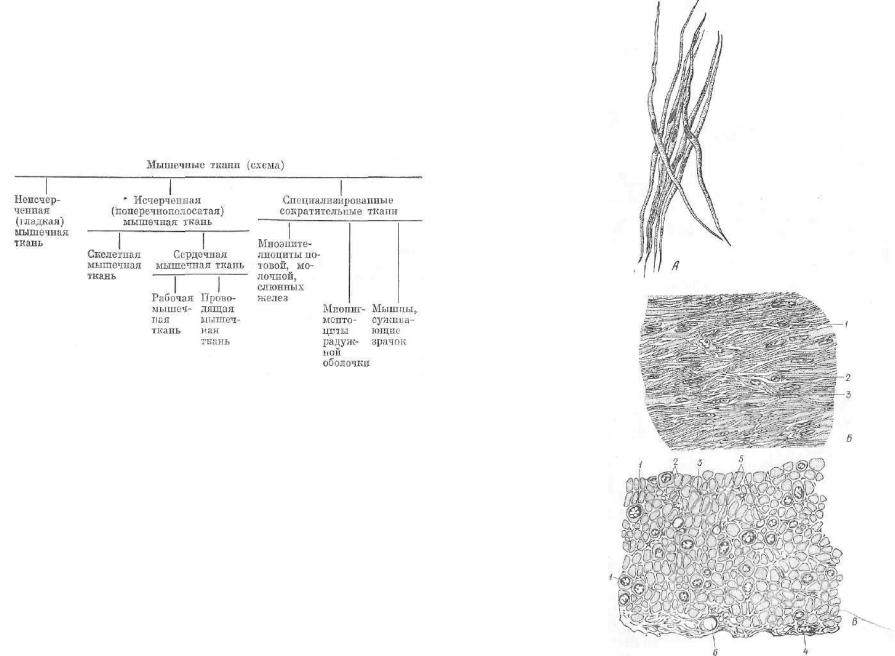
Учитывая особенности строения, место расположения, функ цию и генез, все мышечные ткани можно разделить на три раз новидности: неисчерченная (гладкая), исчерченная (поперечно полосатая), специализированные сократительные ткани эпите лиального и нейроглиального происхождения. Исчерченная! мышечная ткань подразделяется на скелетную и сердечную. Сер дечную мышечную ткань классифицируют на рабочую и прово дящую. К специализированным сократительным тканям относят миоэпителиальные клетки молочной, потовой и слюнных желез;, мышцы, суживающие зрачок; миопигментоциты радужной обо лочки (см. схему).
Мышечные ткани (схема)
НЕИСЧЕРЧЕННАЯ (ГЛАДКАЯ] МЫШЕЧНАЯ ТКАНЬ
Гладкая мышечная ткань относится к ткани с непроизвольным сокращением, так как ее функцию контролирует вегетативная нервная система. Сокращения гладких мышц могут быть медлен ными, но достигать большой силы сжатия. Эта ткань находится в стенках многих полых внутренних органов, поэтому ее именуют внутренностной сократительной тканью. Встречается она в стен ках желудка, кишечника, матки, мочевого пузыря, мочеточников,, бронхов, а также в средних и крупных кровеносных сосудах. Источником развития ее является мезенхима.
Клетки гладкомышечной ткани — миоциты (рис. 134) вытяну той с заостренными концами веретеновидной формы. Их длина варьирует от 10 до 500 мкм, а толщина до 10 мкм. Миоцит — это' одноядерная клетка. Палочковидное ядро занимает в клетке цен тральное положение. Оно содержит распыленный гетерохроматин и одно или два хорошо заметных ядрышка. Клеточный центр (цитоцентр) находится вблизи одной из поверхностей ядра. Ос тальные органеллы сконцентрированы у полюсов палочковид ного ядра. Хорошо развиты митохондрии, комплекс Гольджи, гладкая эндоплазматическая (саркоплазматическая) сеть; незнат
Рис. 134. Гладкомышечные клетки:
А — изолированные; В — в продольном сечении (1— ядро; 2 — ядро фибробласта; 3 — рыхлая соедини тельная ткань); В — в по перечном сечении (I — ядро, 2 — цитоплазма, S— соединительная ткань, 4 — ядро фибробласта, 5 — ка пилляр, в — артерия).
182