

gistologia
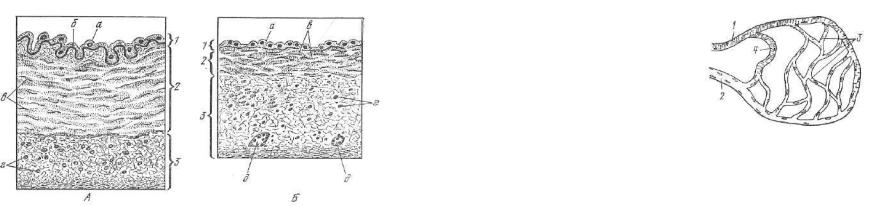
.pdfРпс. 198. Схема строения участка рабочей мышечной ткани миокарда:
1 — миокардиоциты; г — анастомозы; г — вставочные участки; 4 — ядра миокардиоцитов; 5 •— ядра эндотелия капилляров.
ке без участия медиатора. Каждый сердечный миоцит содержит 1 — 2 ядра, расположенные в центре клетки, миофибриллы зани мают периферическую часть цитоплазмы. Между миофибриллами одиночно, группами или цепочками расположены митохондрии, для которых характерно большое количество крист. Миофибрил лы окружены системой трубочек -и канальцев саркоплазматиче-
ской сети. Развита Т-система, |
образованная трубчатыми впячи- |
{ 2 |
' 7 |
1 — сарколемма; 2 — плавмолемма; з — митохондрии; 4 — миофибрилла; 5 — миовиновые филаменты; 6 — актиновые филаменты; 7 — граница между миокардиоцитами; 8 — аона вплетения антиновых миофиламентов; 9 — десмосома; 10 — щеле вой контакт; 11 — канальцы саркоплазматической сети.
264
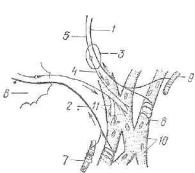
Рис. 200. Схема проводящей системы сердца: |
|
|
|
|
к^щ |
||
1 — синусно-предсердный узел; 2 — предсердно-желу- |
|
|
/? М?<3^> |
||||
дочковый узел; 3 — предсердно-желудочковый ствол |
/ |
|
|
7^-!Ш*Щь^^=^^Ц |
|||
(пучок Гиса); 4 — его ножки и разветвления. |
/" |
|
|
• S ^ ^ ^ T ' ^ ^ f e b j |
|||
ваниями плазмолеммы миоцита, причем Т- |
З-^ШЖ^ИШк ТШ |
||||||
трубочки расположены на уровне Z-поло- |
|
|
|
|
шШ^^ШхлШ |
||
сок миофибрилл. В околоядерной зоне cap- ^'^^ЩзШ^Ш^Ш^ |
|||||||
коплазмы расположены скопления мито- |
|
^ Щ ^ Л Ш Б Ш / |
|||||
хондрий, |
комплекс |
Гольджи, лизосомы, |
|
^Щ===К^? |
|||
гранулы |
гликогена |
и пигмента липофус- |
|
•. |
^Щ^ |
||
цина. |
|
|
|
|
|
|
|
Миоциты окружены тонковолокнистой соединительной тканью |
|||||||
(эпдомизий миокарда), в которой содержатся |
|
|
многочисленные |
||||
капилляры, обеспечивающие процессы микроциркуляции. В серд |
||||||
це имеется о п о р н ы й |
с к е л е т , |
состоящий из фиброзных ко |
||||
лец на границе между |
предсердиями и |
желудочками и в устьях |
||||
сосудов, |
выходящих из желудочков. |
В составе |
фиброзных колец |
|||
находятся пучки плотной коллагеновой |
соединительной ткани, а |
|||||
в сердце животных |
хрящевая и даже |
костная |
ткань (крупный |
|||
рогатый скот). Фиброзно-хрящевые |
кольца препятствуют растя |
|||||
жению |
отверстий и обеспечивают прикрепление свободных кон |
|||||
цов волокон миокарда. |
|
|
|
|
||
А в т о м а т и з м |
сокращения сердца, |
закономерная последова |
||||
тельность сокращений предсердий и желудочков на протяжении |
|
|||||||||||
сердечного цикла |
обусловлены деятельностью |
атипичной мышеч |
||||||||||
ной ткани, входящей в состав п р о в о д я щ е й |
с и с т е м ы серд - |
|
||||||||||
ц а. |
Эта система состоит из |
синусно-предсердяого |
узла, |
располо |
||||||||
женного в устье краниальной полой |
вены, предсердно-желудоч- |
|
||||||||||
кового |
узла, лежащего в межпредсердпой перегородке |
вблизи |
|
|||||||||
места |
прикрепления створки трехстворчатого |
клапана, предсерд |
||||||||||
но-желу дочкового ствола (пучка Гиса) и его разветвлений, рас |
||||||||||||
положенных под эндокардом межжелудочковой перегородки и в |
|
|||||||||||
соединительнотканных прослойках миокарда — волокна Пуркине |
||||||||||||
(рис. 200). Все эти компоненты проводящей системы образованы |
|
|||||||||||
атипичными |
мышечными клетками, |
которые |
в функциональном |
|
||||||||
отношении специализированы или |
на |
выработке |
импульса рас |
|
||||||||
пространяющегося |
по всему |
сердцу и вызывающего сокращение |
|
|||||||||
его отделов в необходимой последовательности и с |
определенной |
|
||||||||||
чавтотой (клетки |
узлов), или на его |
проведении и передаче со |
|
|||||||||
кратительным |
миоцитам. |
|
|
|
|
|
|
|
|
|||
А т и п и ч н ы е |
м и о ц и т ы |
проводящей системы имеют харак |
|
|||||||||
терные |
микроскопические и |
ультраструктурные признаки, отли |
|
|||||||||
чающие их от сократительных миоцитов. При обычной гематоксн- |
|
|||||||||||
линовой окраске они более светлые, имеют неправильно овальную |
|
|||||||||||
форму |
и, как |
правило, поперечный диаметр их больше, чем диа |
|
|||||||||
метр сократительных миоцитов, в 2—3 |
раза. |
Однако в |
составе |
|
||||||||
синусно-предсердного узла обнаружены |
мелкие клетки |
округлой |
|
|||||||||
формы. |
В функциональном |
отношении |
это |
водители |
ритма — |
|
||||||
|
|
|
|
|
|
|
|
|
|
|
265 |
' |
пеисмекеры. Весьма характерным для атипичных миоцитов яв ляются большой объем саркоплазмы и слабое развитие миофибриллярного аппарата. Миофибриллы занимают самую перифери ческую часть в цитоплазме клеток, не имеют параллельной ори ентации, вследствие чего для атипичных миоцитов несвойственна поперечная исчерченность. У них слабо развит саркоплазматический ретикулум, отсутствует система Т-трубок, а в саркоплазме мало митохондрий, но имеется большое количество гранул гли когена. В этих клетках много гликолитических ферментов и уменьшенное количество ферментов аэробного окисления (сукцинатдегидрогеназы и цитохромоксидазы), что свидетельствует о преобладании в них анаэробного гликолиза. Клетки проводящей системы значительно более устойчивы к кислородному голода нию, чем сократительные миоциты.
Эпикард — наружная оболочка сердца. По строению представ ляет тонкую серозную оболочку, состоящую из соединительно тканной основы, содержащей разнообразно ориентированные коллагеновые и эластические волокна, и поверхностного слоя — плоского однослойного эпителия (мезотелия). В соединительной ткани эпикарда проходят крупные кровеносные сосуды и име ется жировая ткань.
Эпикард является висцеральным листком перикарда, плотно срастающимся с миокардом и в области корней крупных сосудов сердца переходящим в париетальный листок. Между этими лист ками имеется щелевидная п е р и к а р д и а л ь н а я п о л о с т ь , содержащая серозную жидкость, выделяемую клетками мезоте лия. Постоянное увлажнение поверхности эпикарда жидкостью уменьшает его трение о париетальный листок перикарда и обес печивает лучшую подвижность сердца.
Кровоснабжение и иннервация сердца. Кровь доставляется к
стенке сердца по правой и левой |
в е н е ч н ы м |
(коронарным) ар - |
т е р и я м , ответвляющимся от |
аорты вблизи |
ее клапана. По |
строению они относятся к артериям мышечно-эластического типа. Венечные артерии разветвляются на ряд мелких артерий, снаб жающих кровью оболочки сердца. Между мелкими ветвями арте
рий и вен имеются а н а с т о м о з ы . В створках |
клапанов сердца |
|
кровеносных сосудов нет. |
В миокарде большое |
количество ка |
п и л л я р о в густой сетью |
оплетают волокна, образуя узкопет |
|
листую сеть, обеспечивающую процессы микроциркуляции. Ка |
||
пиллярные сети вытянуты |
вдоль мышечных волокон. Показано, |
||||||
что каждый сократительный |
миоцит |
находится |
в контакте не |
||||
меньше чем с двумя капиллярами. Кровь из |
капилляров соби |
||||||
рается в к о р о н а р н ы е |
в е н ы , впадающие |
в |
правое пред |
||||
сердие. |
|
|
|
|
|
|
|
Проводящая система, в |
клетках которой |
происходит генера |
|||||
ция возбуждающих |
импульсов, |
способна обеспечить автоматиче |
|||||
ские ритмические |
сокращения |
сердца лишь в покое. В условиях |
|||||
деятельности организма работа сердца |
находится под непрерыв |
||||||
ным воздействием |
нервной |
системы. |
И н н е р в а ц и я сердца |
||||
266
"*.
У Ц Ц Щ Д Ц Д Д Д Д Д Д Д Д WlfHP^MHiH m»
Рис. 201. Схема иннервации сердца:
I — афферентное волокно блуждающего нерва; 2 — афферентное волокно, про ходящее через узел; з — внутрисердечный парасимпатический узел; 4 — постганглионарное волокно; 5 — преганглионарное волокно; 6 — звездчатый сим патический узел; 7 — механорецепторы; 8 — мышечные рецепторы; 9 — крове
носный сосуд; 10 — миокардиоциты; (у II — двигательные нервные окончания.
осуществляется волокнами сим патического и блуждающего
нервов, образующих в оболочках нервные сплетения с интрамуральными ганглиями. В составе постганглионарных симпатических волокон' находятся аксоны клеток звездчатого ганглия и клеток передних грудных симпатических узлов. Концевые утолщения аксонов образуют в сердце двигательные нервные окончания. Парасимпатические волокна содержат аксоны клеток, тела их располагаются в ядре блуждающего нерва в продолговатом мозгу. В сердце опи образуют синапсы на нейронах внутрисердечного ганглия, аксоны которых заканчиваются на мышечных клетках.
Афферентная иннервация осуществляется дендритами нерв ных клеток, тела которых находятся в узловатом ганглии блуж дающего нерва и спинномозговых узлах первых шести грудных сегментов спинного мозга. Концевые веточки дендритов в мио карде формируют многочисленные чувствительные нервные окон
чания, которые можно |
разделить на две группы. Одна группа — |
м е х а н о р е ц е п т о р ы , |
расположенные в соединительнотканных |
прослойках и вокруг артериол. В них возникает сигнал при изме нениях просвета кровеносных сосудов и растяжении соединитель ной ткани. Центростремительные импульсы от этих рецепторов
вызывают рефлекторное |
ускорение |
ритма сердца. |
Другая груп |
|
п а — м ц ш е ч н ы е |
р е ц е п т о р ы , |
имеющие вид |
спирали. Они |
|
специализированы |
для |
сигнализации о сокращении миоцитов |
||
(рис. 201). Кроме того, с участием |
различных нервных клеток, |
|||
сосредоточенных в интракардиальиых ганглиях, образуются мест ные рефлекторные дуги.
КРОВЕНОСНЫЕ СОСУДЫ
Кровеносные сосуды представляют замкнутую систему развет
вленных |
трубок разного диаметра, входящих в состав большого |
и малого |
кругов кровообращения. В этой' системе различают: |
а р т е р и и , по которым кровь течет от сердца к органам и тка ням, в е н ы — по ним кровь возвращается в сердце, и комплекс сосудов м и к р о ц и р к у л я т о р н о г о р у с л а , обеспечивающих наряду с транспортной функцией обмен веществ между кровью и окружающими тканями.
267
Кровеносные сосуды р а з в и в а ю т с я из мезенхимы. В эм бриогенезе наиболее ранний период характеризуется появлением
многочисленных клеточных скоплений |
мезенхимы в стенке жел |
||||||||||
точного |
мешка — кровяных островков. |
Внутри |
островка образу |
||||||||
ются |
кровяные клетки и формируется полость, а располоя^енные |
||||||||||
но периферии |
клетки |
становятся |
плоскими, соединяются между |
||||||||
собой при помощи |
клеточных контактов и формируют эндотели- |
||||||||||
альную |
выстилку |
образующейся |
трубочки. |
Такие |
первичные |
||||||
кровеносные трубочки по мере |
образования |
соединяются между |
|||||||||
собой и формируют капиллярную |
сеть. Окружающие клетки ме |
||||||||||
зенхимы |
превращаются в перициты, гладкие |
мышечные клетки |
|||||||||
и адвентициальные клеткщ В теле зародыша |
кровеносные капил |
||||||||||
ляры |
закладываются из клеток |
мезенхимы |
вокруг |
щелевидных |
|||||||
пространств, заполненных тканевой жидкостью. |
Когда по сосу |
||||||||||
дам усиливается кровоток, эти клетки |
становятся |
эндотелиаль- |
|||||||||
ными, а из окружающей мезенхимы |
формируются элементы |
||||||||||
средней |
и наружной оболочки. |
|
|
|
|
|
|
||||
Сосудистая |
система |
обладает |
очень |
большой п л а с т и ч н о |
|||||||
стью . |
Прежде |
всего отмечается |
значительная |
изменчивость гу |
|||||||
стоты |
сосудистой сети, |
так как в |
зависимости |
от |
потребностей |
||||||
органа в питательных веществах и кислороде в широких пределах колеблется количество приносимой ему крови. Изменение скоро сти кровотока и кровяного давления ведет к образованию новых сосудов и перестройке имеющихся сосудов. Происходит превра щение мелкого сосуда в более крупный с характерными особен ностями строения его стенки. Наибольшие изменения возникают в сосудистой системе при развитии окольного, или коллатераль ного, кровообращения.
Артерии и вены построены по единому плану — в их стенках различают три оболочки: внутреннюю (tunica intima), среднюю (tunica media) и наружную (tunica adventicia). Однако степень развития этих оболочек, их толщина и тканевый состав тесно связаны с функцией, выполняемой сосудом и гемодинамическими условиями (высотой кровяного давления и скоростью крово тока), которые в различных отделах сосудистого русла неоди наковы.
А р т е р и и . По строению |
стенок различают артерии мышеч |
ного, мышечно-эластического |
и эластического типов. |
К артериям эластического типа относятся аорта и легочная артерия. В соответствии с высоким гидростатическим давлением (до 200 мм ртутного столба), создаваемым нагнетательной дея тельностью желудочков сердца, и большой скоростью кровотока (0,5—1 м/с) у этих сосудов резко выражены упругие свойства, которые обеспечивают прочность стенки при ее растяжении и воз вращении в исходное положение, а также способствуют превра щению пульсирующего кровотока в постоянный непрерывный. Стенка артерий эластического типа отличается значительной толщиной и наличием большого количества эластических элемен тов в составе всех оболочек.
В н у т р е н н я я о б о л о ч к а |
состоит из двух слоев — эндоте- |
|
яиального и подэндотелиального. |
Эндотелиальные клетки, форми |
|
рующие |
сплошную внутреннюю |
выстилку, имеют различную ве |
личину |
и форму, содержат одно или несколько ядер. В их цито |
|
плазме немногочисленные органеллы и много микрофиламентов. Под эндотелием находится базальная мембрана. Подэндотелиальный слой состоит из рыхлой тонковолокнистой соединительной ткани, в составе которой наряду с сетью эластических волокон присутствуют малодифференцированные клетки звездчатой фор мы, макрофаги, гладкие мышечные клетки. В аморфном веществе этого слоя, имеющем большое значение для питания стенки, со держится значительное количество гликозаминогликанов. При повреждении стенки и развитии патологического процесса (ате росклерозе) в подэндотелиальном слое накапливаются липиды (холестерин и его эфиры). Клеточные элементы подэндотелиаль ного слоя играют важную роль в регенерации стенки. На границе
со средней |
оболочкой |
располагается |
густая сеть эластических |
||||
волокон. |
|
|
|
|
|
|
|
С р е д н я я |
о б о л о ч к а |
состоит из многочисленных эластиче |
|||||
ских окончатых |
мембран, |
между которыми располагаются косо |
|||||
ориентированные пучки гладких мышечных клеток. Через |
окна, |
||||||
(фенестры) |
мембран |
осуществляется |
внутристеночный |
транс |
|||
порт веществ, необходимых для |
питания клеток стенки. Как мем |
||||||
браны, так |
и клетки |
гладкой |
мышечной ткани окружены |
сетью |
|||
эластических волокон, |
формирующих |
вместе с волокнами |
внут |
||||
ренней и наружной оболочек единый каркас, обеспечивающий: высокую эластичность стенки.
Н а р у ж н а я |
о б о л о ч к а |
образована |
соединительной тка |
нью, в которой |
преобладают пучки коллагеновых волокон, ори |
||
ентированных продольно. В этой оболочке |
расположены и вет |
||
вятся сосуды, обеспечивающие |
питание как наружной оболочки, |
||
так и наружных |
зон средней оболочки. |
|
|
Артерии мышечного типа. К разным по калибру артериям этого типа относится большинство артерий, доставляющих и ре гулирующих приток крови к различным частям и органам орга низма (плечевая, бедреппая, селезеночная и др.). При микроско пическом исследовании в стенке хорошо различимы элементы всех трех оболочек (рис. 202).
В н у т р е н н я я о б о л о ч к |
а |
состоит из |
трех слоев: эндоте- |
лиального, подэндотелиального |
и |
внутренней |
эластической мем |
браны. Эндотелий имеет вид тонкой пластинки, состоящей из
вытянутых вдоль сосуда клеток с овальными, |
выступающими в |
|||||
просвет ядрами. |
Подэндотелиальный слой более |
развит в круп |
||||
ных по |
диаметру |
артериях и состоит из клеток |
звездчатой или |
|||
веретенообразной |
формы, тонких эластических |
волокон и аморф |
||||
ного вещества, содержащего |
гликозаминогликаны. На границе со |
|||||
средней |
оболочкой лежит |
в н у т р е н н я я |
э л а с т и ч е с к а я |
|||
м е м б р а н а , |
хорошо заметная на препаратах в виде блестящей, |
|||||
окрашенной |
эозином в светло-розовый цвет волнистой полоски. |
|||||
268 |
269 |
|
Эта мембрана пронизана многочисленными отверетиями, имею щими значение для транспорта веществ.
С р е д н я я о б о л о ч к а построена преимущественно из глад кой мышечной ткани, пучки клеток которой идут по спирали,, однако при изменении положения артериальной стенки (растя жении) расположение мышечных клеток может изменяться. Со кращение мышечной ткани средней оболочки имеет значение в регулировании притока крови к органам и тканям в соответствии с их потребностями и поддержании кровяного давления. Между пучками клеток мышечной ткани расположена сеть эластичен ских волокон, которые вместе с эластическими волокнами подэндотелиального слоя и наружной оболочки формируют единый эластический каркас, придающий стенке упругость при ее сдав ливании. На границе с наружной оболочкой в крупных артериях
мышечного типа |
имеется |
н а р у ж н а я |
э л а с т и ч е с к а я |
мем |
|||||
б р а н а , |
состоящая из |
плотного сплетения продольно |
ориенти |
||||||
рованных |
эластических |
волокон. |
В более |
мелких артериях эта |
|||||
мембрана |
не выражена. |
|
|
|
|
|
|
|
|
Н а р у ж н а я |
о б о л о ч к а состоит |
из |
соединительной |
ткани,. |
|||||
в которой коллагеновые |
|
волокна |
и сети |
эластических |
волокон |
||||
вытянуты в продольном |
направлении. Между волокнами |
распо |
|||||||
лагаются |
клетки, преимущественно |
фиброциты. В наружной обо |
|||||||
лочке находятся |
нервные |
волокна и мелкие кровеносные |
сосу |
||||||
ды, питающие наружные слои стенки артерии. |
|
|
|||||||
Артерии мышечно-эластического типа по строению стенки за нимают промежуточное положение между артериями эластиче ского и мышечного типа. В средней оболочке в равном количе стве развиты спирально ориентированная гладкая мышечная! , ткань, эластические пластины и сеть эластических волокон.
270
Рис. 203. Схема сосудов микроциркуляторного русла: \
1 — артериола; 2 — венула; з — капиллярная сеть; 4 — артерирло-венулярный анастомоз.
С о с у д ы |
м и к р о ц и р к у л я - |
т о р н о г о р у с л а . |
На месте перехо |
да артериального русла в венозное в |
|
органах и тканях сформирована густая сеть мелких прекапиллярных, капиллярных и посткапиллярных сосудов. Этот комплекс мел ких сосудов, обеспечивающий кровенаполнение органов, транссо судистый обмен и тканевый гомеостаз, объединяют термином микроциркуляторное русло. В его состав входят различные артериолы, капилляры, венулы и артериоло-венулярные анастомозы (рис. 203).
Артериолы, По мере уменьшения диаметра в артериях мы шечного типа истончаются все оболочки и они переходят в арте риолы — сосуды диаметром менее 100 мкм. Внутренняя оболочка
их состоит из эндотелия, расположенного |
на |
базальпой мембра |
|||||||
не, и отдельных клеток подэндотелиального слоя. |
В |
некоторых |
|||||||
артериолах может быть очень тонкая |
внутренняя |
эластическая |
|||||||
мембрана. В |
средней оболочке сохраняется |
один |
ряд спирально |
||||||
расположенных клеток гладкой мышечной ткани. В |
стенке ко |
||||||||
нечных артериол, от которых ответвляются капилляры, |
гладко- |
||||||||
мышечные клетки не образуют сплошного |
ряда, а расположены |
||||||||
разрозненно. Это |
п р е к а п и л л я р н ы е |
а р т е р и о л ы . |
Однако |
||||||
в месте ответвления от артериолы капилляр окружен |
значитель |
||||||||
ным количеством |
гладкомышечных |
клеток, |
которые |
образуют |
|||||
своеобразный |
п р е к а п и л л я р н ы й |
с ф и н к т е р . |
Вследствие |
||||||
изменения тонуса таких сфинктеров регулируется |
кровоток в ка |
||||||||
пиллярах соответствующего участка |
ткани |
или органа. |
Между |
||||||
мышечными клетками имеются эластические волокна. Наружная
оболочка |
содержит отдельные |
адвентициальные клетки и кол |
||||
лагеновые |
волокна. |
|
|
|
||
Капилляры — важнейшие элементы микроциркуляторного рус |
||||||
ла, в которых осуществляется |
обмен газами и различными веще |
|||||
ствами |
между |
кровью и окружающими тканями. В |
большинстве |
|||
органов |
между |
артериолами и венулами |
образуются |
ветвящиеся |
||
к а п и л л я р н ы е с е т и , расположенные |
в рыхлой |
соединитель |
||||
ной ткани. Плотность капиллярной сети в разных органах может
быть различной. |
Чем интенсивнее |
обмен веществ в |
органе, тем |
||
гуще сеть его капилляров. |
Наиболее развита |
сеть |
капилляров |
||
в сером веществе |
органов |
нервной |
системы, в |
органах внутрен |
|
ней секреции, миокарде сердца, вокруг легочных альвеол. В ске летных мышцах, сухожилиях, нервных стволах капиллярные сети ориентированы продольно.
Капиллярная сеть постоянно находится в состоянии пере стройки. В органах и тканях значительное количество капилля ров не функционирует. В их сильно уменьшенной полости цирку-
271
лирует только плазма крови ( п л а з м е н н ы е к а п и л л я р ы)-. Количество открытых капилляров увеличивается при интенсифи кации работы органа.
Капиллярные сети встречаются и между одноименными сосу дами, например венозные капиллярные сети в дольках печени. аденогипофизе, артериальные — в почечных клубочках. Кроме об разования разветвленных сетей, капилляры могут иметь форму капиллярной петли (в сосочковом слое дермы) или формпровать клубочки (сосудистые клубочки почек).
Капилляры — наиболее узкие сосудистые трубочки. Их калибр в среднем соответствует диаметру эритроцита (7—8 мкм), одна ко в зависимости от функционального состояния и органной спе циализации диаметр капилляров может быть различным. Узкие капилляры (диаметром 4—5 мкм) в миокарде. Особые синусоидные капилляры с широким просветом (30 мкм и более) в дольках печени, селезенке, красном костном мозге, органах внутренней секреции.
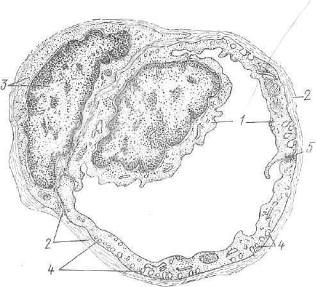
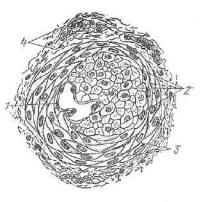
Стенка кровеносных капилляров состоит из нескольких струк турных элементов. Внутреннюю выстилку формирует слой эндо телиальных клеток, расположенных на базальной мембране, в по следней содержатся клетки — перициты. Вокруг базальной мем браны располагаются адвентициальные клетки и ретикулярные волокна (рис. 204).
272
Плоские э н д о т е л и а л ь н ы е к л е т к и вытянуты по длине* капилляра и имеют очень тонкие (менее 0,1 мкм) периферические безъядерные участки. Поэтому при световой микроскопии попе речного среза\ сосуда различима только область расположения ядра толщиной 3—5 мкм. Ядра эндотелиоцитов чаще овальной формы, содержат конденсированный хроматин, сосредоточенный около ядерной оболочки, которая, как правило, имеет неровные контуры. В цитоплазме основная масса органелл расположена в околоядерной области. Внутренняя поверхность эндотелиальных клеток неровная, плазмолемма образует различные по форме и высоте микроворсинки, выступы и клапанообразные структуры,. Последние особенно характерны для венозного отдела капилля ров. Вдоль внутренней и наружной поверхностей эндотелиоцитов
располагаются многочисленные |
п и н о ц и т о з н ы е |
п у з ы р ь к и , , |
||
свидетельствующие об |
интенсивном поглощении |
и переносе ве |
||
ществ через цитоплазму этих |
клеток. |
Эндотелиальные клетки |
||
благодаря способности |
быстро |
набухать |
и затем, |
отдавая жид |
кость, уменьшаться по высоте могут изменять величину просвета капилляра, что, в свою очередь, влияет на прохождение через него= форменных элементов крови. Кроме того, при электронной мик
роскопии в цитоплазме |
выявлены |
микрофиламенты, |
обусловли |
||
вающие сократительные |
свойства |
эндотелиоцитов. |
|
||
Б а з а л ь н а я |
м е м б р а н а , |
расположенная под |
эндотелием,, |
||
выявляется при |
электронной |
микроскопии и представляет пла |
|||
стинку толщиной 30—35 нм, состоящую из сети тонких фибрилл,- содержащих коллаген IV типа и аморфного компонента. В по следнем наряду с белками содержится гиалуроновая кислота, полимеризованное или деполимеризованное состояние которой обусловливает избирательную проницаемость капилляров. Базаль ная мембрана обеспечивает также эластичность и прочность ка пилляров. В расщеплениях базальной мембраны встречаются осо бые отросчатые клетки — перициты. Они своими отростками охватывают капилляр и, проникая через базальную мембрану, формируют контакты с эндотелиоцитами.
В соответствия с особенностями строения эндотелиальной выстилки и базальной мембраны различают три типа капилля ров. Большинство капилляров в оргапах и тканях принадлежит к первому типу ( к а п и л л я р ы о б щ е г о т и п а ) . Они характе ризуются наличием непрерывных эндотелиальной выстилки и ба зальной мембраны. В этом сплошном слое плазмолеммы сосед них эндотелиальных клеток максимально сближены и образуют соединения по типу плотного контакта, который непроницаем для макромолекул. Встречаются и другие виды контактов, когда края соседних клеток налегают друг на друга наподобие черепицы или соединяются зубчатыми поверхностями. По длине капилляров выделяют более узкую (5—7 мкм) проксимальную (артериолярную) и более широкую (8—10 мкм) дистальную (венулярную) части. В полости проксимальной части гидростатическое давле ние больше коллоидно-осмотического, создаваемого находящими-
18 Заказ № 908 |
273 |
ися в крови белками. В результате жидкость фильтруется за стенжу. В дистальной части гидростатическое давление становится меньше коллоидно-осмотического, что обусловливает переход во-
.ды и растворенных в ней веществ из окружающей/тканевой жид кости в кровь. Однако выходной поток жидкости больше вход ного, и избыточная жидкость в качестве составной части тканевой жидкости соединительной тка<ни поступает в лимфатическую «систему.
В некоторых органах, в которых интенсивно происходят про цессы всасывания и выделения жидкости, а также быстрый транс порт в кровь макромолекулярных веществ, эндотелий капилляров имеет округлые субмикроскопические отверстия диаметром 60— 80 нм или округлые участки, затянутые тонкой диафрагмой (поч ки, органы внутренней секреции). Это к а п и л л я р ы с ф е н е - •с т р а м и (лат. fenestrae — окна).
Капилляры третьего |
типа — с и н у с о и д н ы е , характеризу |
ются большим диаметром |
своего просвета, наличием между эндо- |
телиальными клетками |
широких щелей и прерывистой базаль- |
пой мембраной. Капилляры этого типа обнаружены в селезенке, зсрасном костном мозге. Через их стенки проникают не только макромолекулы, но и клетки крови.
Венулы — отводящий отдел микроциркуляторного русла и на чальное звено венозного отдела сосудистой системы. В них со бирается кровь из капиллярного русла. Диаметр их просвета бо лее широкий, чем в капиллярах (15—50 мкм). В стенке венул, так же как и у капилляров, имеется слой эндотелиальных кле ток, расположенных .на базальной мембране, а также более вы раженная наружная соединительнотканная оболочка. В стенках
венул, переходящих в мелкие |
вены, находятся |
отдельные |
глад |
|
кие мышечные клетки. |
В п о с т к а п и л л я р н ы х в е н у л а х |
|||
•тимуса, лимфатических |
узлов |
эндотелиальная |
выстилка |
пред |
ставлена высокими эндотелиальными клетками, способствующи ми избирательной миграции лимфоцитов при их рециркуляции. В венулах вследствие тонкости их стенки, медленного кровотока и низкого кровяного давления может депонироваться значитель ное количество крови.
Артериоло-венулярные анастомозы. Во всех органах обнару жены трубочки, по которым кровь из артериол может направ ляться непосредственно в венулы, минуя капиллярную сеть. Особенно много анастомозов в дерме кожи, в ушной раковине, гребне птиц, где играют определенную роль в терморегуляции.
По строению истинные артериоло-венулярные анастомозы {шунты) характеризуются наличием в стенке значительного ко личества продольно ориентированных пучков из гладких мышеч ных клеток, расположенных или в подэндотелиальном слое инти мы (рис. 205), или во внутренней зоне средней оболочки. В не которых анастомозах эти клетки приобретают эпителиоподобный вид. Продольно расположенные мышечные клетки находятся и © наружной оболочке. Встречаются не только простые аыастомо-
274 |
18* |
275. |
ратному течению. Число клапанов больше в тех венах, в которых кровь течет в направлении, обратном действию силы тяжести (на пример, в венах конечностей).
По степени развития в стенке мышечных элементов разли чают вены безмышечного и мышечного типов.
Вены безмышечного типа. К характерным венам данного ти па относят вены костей, центральные вены печеночных долек и трабекулярные вены селезенки. Стенка этих вен состоит только из слоя эпдотелиальных клеток, расположенных на базальной мембране, и наружного топкого слоя волокнистой соединительной ткани. С участием последней стенка плотно срастается с окру жающими тканями, вследствие чего эти вены пассивны в про движении по ним крови и не спадаются. Безмышечные вены мозговых оболочек и сетчатки глаза, наполняясь кровью, способ ны легко растягиваться, но в то же время кровь под действием собственной силы тяжести легко оттекает в более крупные веноз ные стволы.
Вены мышечного типа. Стенка этих вен, подобно стенке ар терий, состоит из трех оболочек, однако границы между ними ме нее отчетливы. Толщина мышечной оболочки в стенке вен разной локализации неодинаковая, что зависит от того, движется кровь в них под действием силы тяжести или против нее. На основании этого вены мышечного типа подразделяют на вены со слабым, средним и сильным развитием мышечных элементов. К венам первой разновидности относят горизонтально расположенные вены верхней части туловища организма и вены пищеваритель ного тракта. Стенки таких вен тонкие, в их средней оболочке гладкая мышечная ткань не образует сплошного слоя, а располо жена пучками, между которыми имеются прослойки рыхлой со единительной ткани.
К венам с сильным развитием мышечных элементов относят крупные вены конечностей животных, по которым кровь течет вверх, против силы тяжести (бедренная, плечевая и др.). Для них характерны продольно расположенные небольшие пучки клеток гладкой мышечной ткани в подэндотелиальном слое интимы и хорошо развитые пучки этой ткани в наружной оболочке. Сокра щение гладкой мышечной ткани наружной и внутренней оболо чек приводит к образованию поперечных складок стенки вей, что препятствует обратному кровотоку.
В средней оболочке содержатся циркулярно расположенные пучки клеток гладкой мышечной ткани, сокращения которых способствуют продвижению крови к сердцу. В венах конечностей имеются клапаны, представляющие собой тонкие складки, обра зованные эндотелием и подэндотелиальным слоем. Основу кла пана составляет волокнистая соединительная ткань, которая в основании створок клапана может содержать некоторое количе ство клеток гладкой мышечной ткани. Клапаны также препят ствуют обратному току венозной крови. Для движения крови в венах существенное значение имеют присасывающее действие
276
грудной клетки во время вдоха и сокращение скелетной мышеч ной ткани, окружающей венозные сосуды.
В а с к у л я р из а ц и я и и н н е р в а ц и я к р о в е н о с н ы х с о с у д о в . Питание стенки крупных и средних артериальных со
судов осуществляется как извне — через |
сосуды сосудов (vasa |
|
vasorum), так и изнутри — за |
счет крови, |
протекающей внутри |
сосуда. Сосуды сосудов — это |
ветви тонких околососудистых арте |
|
рий, проходящих в окружающей соединительной ткани. В наруж ной оболочке стенки сосуда ветвятся артериальные веточки, в среднюю проникают капилляры, кровь из которых собирается в венозные сосуды сосудов. Интима и внутренняя зона средней оболочки артерий не имеют капилляров и питаются со стороны просвета сосудов. В связи со значительно меньшей силой пуль совой волны, меньшей толщиной средней оболочки, отсутствием внутренней эластической мембраны механизм питания вены со стороны полости не имеет особого значения. В венах сосуды со судов снабжают артериальной кровью все три оболочки.
Сужение и расширение кровеносных сосудов, поддержание сосудистого тонуса происходят главным образом под влиянием импульсов, поступающих из сосудодвигательного центра. Импульсы от центра передаются к клеткам боковых рогов спинного мозга, откуда к сосудам поступают по симпатическим нервным волок нам. Конечные разветвления симпатических волокон, в составе которых находятся аксоны нервных клеток симпатических ганг лиев, образуют на клетках гладкой мышечной ткани двигательпые нервные окончания. Эфферентная симпатическая иннерва ция сосудистой стенки обусловливает основной сосудосуживаю щий эффект. Вопрос о природе вазодилататоров окончательно не решен.
Установлено, что сосудорасширяющими в отношении сосудов головы являются парасимпатические нервные волокна.
Во всех |
трех оболочках стенки |
сосудов концевые |
разветвле |
ния дендритов нервных клеток, |
преимущественно спинальных |
||
ганглиев, |
образуют многочисленные чувствительные |
нервные |
|
окончания. |
В адвентиции и околососудистой рыхлой соединитель |
||
ной ткани |
среди многообразных по форме свободных |
окончаний |
|
встречаются и инкапсулированные тельца. Особенно важное фи зиологическое значение имеют специализированные интерорецепторы, воспринимающие изменения давления крови и ее химиче ского состава, сосредоточенные в стенке дуги аорты и в области
разветвления сонной |
артерии |
на |
внутреннюю и |
наружную — |
|||||
аортальная |
и |
каротидная |
рефлексогенные |
зоны. |
Установлено, |
||||
что помимо |
этих |
зон |
существует достаточное количество других |
||||||
сосудистых |
территорий, чувствительных к |
изменению давления |
|||||||
и химического |
состава |
крови (баро- и хеморецепторы). От рецеп |
|||||||
торов всех |
специализированных территорий импульсы по цен |
||||||||
тростремительным |
нервам |
достигают |
сосудодвигательного центра |
||||||
продолговатого |
мозга, |
вызывая |
соответствующую |
компенсатор |
|||||
ную нервнорефлекторную |
реакцию. |
|
|
|
|||||
|
|
|
|
|
|
|
|
|
277 |
В состав системы лимфатических сосудов входят элементы микролимфоносного русла — лимфатические капилляры и посткапил ляры, внутри- и внеорганные лимфатические сосуды, отводящие лимфу от органов, и главные лимфатические стволы — грудной проток и правый лимфатический проток, впадающие в краниаль ную полую вену. Лимфатические сосуды образуют систему отто ка лимфы, в которой она движется в одном направлении. Посред ством лимфатических капилляров и сосудов из тканей удаляются продукты обмена веществ, крупномолекулярные вещества, мик роорганизмы и другие частицы. По лимфатическим сосудам про исходит рециркуляция в организме лимфоцитов. По ним могут распространяться клетки злокачественных опухолей.
Л и м ф а т и ч е с |
к и е |
к а п и л л я р ы |
представляют собой |
замкнутые с одного |
конца |
тонкостенные |
трубочки, которые, раз |
ветвляясь и соединяясь, формируют в органах крупнопетлистую сеть. Наиболее густые лимфокапиллярные сети в подкожной клетчатке, в оболочках внутренних органов и капсуле суставов. Лимфатические капилляры отсутствуют в головном и спинном мозге, костях, гиалиновом хряще, роговице и хрусталике глаза. Диаметр лимфатических капилляров различный, но, как прави ло, в несколько раз превышает диаметр кровеносных капилляров. Стенка лимфокапилляров состоит только из уплощенных эндотелиальных клеток, к наружной поверхности которых прикрепля ются особые «якорные» микрофиламенты, благодаря которым обеспечивается прочное прикрепление эндотелиальной трубки к коллагеновым фибриллам окружающей соединительной ткани. Базальная мембрана отсутствует. Между эндотелиальными клет ками имеются щелевидные пространства, через которые в полость
капилляров |
проникают крупномолекулярные вещества, |
частицы |
и клетки. |
В цитоплазме эндотелиоцитов обнаруживают |
актино- |
вые микрофиламенты, сокращение которых регулирует прони цаемость стенки.
Лимфатические сосуды в зависимости от калибра делят на мелкие, средние и крупные. Отличительной особенностью строе ния их стенки является наличие клапанов, позволяющих лимфе продвигаться лишь в одном направлении — к сердцу. В переме щении лимфы определенное значение имеют сокращение мышеч ной ткани стенки самого сосуда и окружающих скелетных мышц, пульсовые волны соседних артерий и другие факторы. В местах расположения клапанов лимфатические сосуды при наполнении
лимфой колбовидно |
расширяются. |
М е л к и е в н у т р и о р г а н - |
||
н ы е л и м ф а т и ч е с к и е с о с у д ы |
по |
диаметру нередко уже, |
||
чем лимфатические капилляры. Это |
эндотелиальные трубки, ок |
|||
руженные |
соединительнотканной оболочкой, в которой по мере |
|||
увеличения |
калибра |
могут встречаться |
гладкие мышечные |
|
клетки. |
|
|
|
|
278 |
|
|
|
|
Г Л А В А 7
•ОРГАНЫ ГЕМОПОЭЗА И ИММУНОЛОГИЧЕСКОЙ ЗАЩИТЫ
В системе органов гемопоэза (кроветворения) и иммунологиче ской защиты различают центральные и периферические органы. К центральным органам у животных относят красный костный мозг и тимус, а у птиц также и фабрициеву сумку. В период эм бриогенеза эти органы заселяются полипотентными стволовыми кроветворными клетками, обладающими свойством самоподдер жания в течение всей жизни организма. Из них в красном кост ном мозге образуются эритроциты, гранулоциты, моноциты и кровяные пластинки. В условиях специфического микроокруже ния, создаваемого соответствующими клетками стромы и макро фагами, в центральных органах происходит антигеннезависимое развитие многообразных клеток-предшественников и образование иммунокомпетентных Т- и В-лимфоцитов, которые выходят в кро вяное русло и в дальнейшем заселяют определенные зоны в пе риферических органах.
К периферическим органам гемопоэза относят лимфатические узлы, селезенку и лимфоидные образования стенки пищевари тельного тракта, а у птиц также кожи и легких. В лимфоидной ткани этих органов и образований происходит зависимый от ан тигенов процесс пролиферации Т- и В-лимфоцитов и превраще ние их в эффекторные клетки, обеспечивающие различные реак ции клеточного и гуморального иммунитета. Таким образом, лимфоидная ткань периферических органов кроветворения бла годаря непрерывно происходящим в ней процессам размножения, миграции и рецпркуляции лимфоцитов, несмотря на топографи-
279
ческую разобщенность, представляет собой единую защитную систему организма.
Общий морфологический признак строения всех кроветворных органов — наличие в их составе отростчатых клеток ретикулярной ткани мезенхимного происхождения (красный костный мозг,. лимфатические узлы, селезенка, лимфоидная ткань слизистых оболочек) или сети отростчатых эпителиальных клеток (тимус.. фабрициева сумка). Клетки ретикулярной или ретикулоэпителиальной стромы вместе с макрофагами образуют в кроветворных органах специфическую микросреду, в которой создаются опти мальные условия для обеспечения взаимного контакта разнооб разных клеток при их функционировании, а также условия для размножения, созревания и разрушения клеток. Особая система синусоидных капилляров и посткапиллярных венул способствует миграции клеток, создает замедленный внутриорганный ток кро ви и лимфы и их депонирование.
КОСТНЫЙ МОЗГ
Ксстный мозг представляет содержимое полужидкой консистен ции, заполняющее полости костей позвоночных животных. Раз личают красный и желтый костный мозг.
Красный костный мозг (medulla ossium rubra) у взрослых жи вотных находится в плоских костях (ребрах, костях черепа, таза,, грудной кости), позвонках и эпифизах трубчатых костей.
Образование красного костного мозга |
у животных начинается, |
в ранний период эмбриогенеза, во время |
образования хрящевого* |
скелета. У 7-недельного эмбриона овцы в хрящевом скелете ко нечностей в области диафиза появляется полость, в которой раз вивается вначале костный мозг, состоящий из остеобластов,. остеокластов и ретикулярных клеток. Затем вокруг формирую щихся сосудов микроциркуляторного русла происходит усилен ное образование гемопоэтических клеток, входящих в состав миелоидной ткани. К моменту рождения все костные полости запол нены красным костным мозгом, сохраняющимся у грызунов BS во взрослом состоянии. У большинства животных в костях конеч ностей и хвостовых позвонках красный костный мозг превраща ется в желтый (жировой). У птиц он в незначительном количе стве сохраняется во всех костях.
Красный костный мозг является центральным органом кро ветворения, в котором находится самоподдерживающаяся попу ляция стволовых клеток, а также образуются эритроциты, зер нистые лейкоциты, моноциты, В-лимфоциты (у млекопитающих). и кровяные пластинки.
Остовом костного мозга служат анастомозирующие между со бой соединительнотканные перекладины, отходящие от эядоостакости. В пространствах между перекладинами расположена ре тикулярная ткань, пронизанная множеством сосудов микроцир куляторного русла. Среди них важнейшее значение имеют сину-
соидные капилляры, обеспечивающие избирательную миграцию зрелых форменных элементов крови в сосудистое русло. Синусои ды имеют широкий диаметр просвета и многочисленные поры в стенке. В эндотелиальной выстилке этих капилляров и среди ретикулярных клеток находятся макрофаги.
У взрослого животного красный костный мозг характеризует ся полиморфным клеточным составом, меняющимся при различ ных физиологических и патологических состояниях. В ячеях ретикулярной стромы и около синусоидов выделяют группы созре вающих и зрелых свободных гемопоэтических элементов, принад лежащих преимущественно к трем системам клеточного обнов ления — эритропоэтической, гранулопоэтической и мегакариоци- тарно-тромбопоэтической. Среди них встречаются характерные скопления клеток эритроцитарного ряда (эритробласты, базофильные, полихроматофильные и оксифильные нормоциты) вокруг макрофага — эритробластические островки. Центральный макро фаг такого островка переносит накапливаемое железо в разви вающиеся эритроциты, поглощает ядра нормоцитов и фагоцити рует стареющие и погибающие эритроциты. Развивающиеся клетки гранулоцитарного ряда (нейтрофильные, эозинофильные, базофильные миелоциты, промиелоциты и метамиелоциты) также расположены островками, но не связаны с макрофагами. Их лег ко идентифицировать по характерной специфической зернистости в цитоплазме. Костно-мозговой резерв гранулоцитов в 10 раз превышает их число в сосудистой крови.
Среди клеток миелоидной ткани своей гигантской величиной, многодольчатым ядром и неровными контурами выделяются мегакариоциты и мегакариобласты. Они, как правило, расположе ны в контакте со стенкой синусоида, что обеспечивает поступ ление кровяных пластинок непосредственно в кровяное русло. Труднее заметны группы костно-мозговых лимфоцитов (В-лим- фоциты, нулевые лимфоциты), а также лимфоцитоподобные ство ловые клетки и полустволовые предшественники развивающихся клеток и моноциты.
Относительное содержание в красном костном мозге созре вающих и зрелых клеточных элементов каждого из ростков кро ветворения является важным показателем процесса кроветворе ния. В обычных физиологических условиях из костного мозга в сосудистую кровь проникают только зрелые эритроциты и лейко циты, а также некоторое количество стволовых предшественни ков, способных переселяться в другие органы (тимус, фабрициеву сумку — у птиц). Появление незрелых клеток (нормоцитов, миелоцитов) в циркулирующей крови является признаком пато логического состояния.
Одни из постоянных элементов красного костного мозга (бо лее 10%) —жировые клетки. Липоциты костного мозга содержат нейтральные жиры, в состав которых не входит олеиновая кис лота. Голодание не приводит к исчезновению жировых включе ний из липоцитов.
280 |
281 |
Красный костный мозг без резкой границы в области диафизов трубчатых костей переходит в желтый костный мозг, состоя щий из жировой ткани. При увеличении иотребности организма в клетках крови желтый костный мозг может замещаться крас ным (после кровопотерь, при отравлениях ж т. п..).
ТИМУС (ВИЛОЧКОВАЯ ЖЕЛЕЗА)
У большинства животных тимус (thymus) состоит из парных шейных частей, расположенных по бокам трахеи, и непарной ча сти, расположенной в грудной полости. Принадлежит тиму© к центральным органам иммунной системы, контролирующим ее формирование и полноценное функционирование. Свою регуляторную иммуногенную функцию тимус осуществляет посредст вом создания разнородной популяции Т-лимфоцитов, имеющих важнейшее значение в развитии как клеточного, так и гумораль ного иммунитета. Регулирующая функция тимуса связана и с выработкой гуморальных факторов (тимозин и др.), обладающих дистантным действием и воздействующих на лимфоциты в пери ферических лимфоидных органах (лимфатических узлах, селе зенке).
В эмбриогенезе тимус развивается и начинает функциониро вать ранее других лимфоидных органов и образований. У живот ных он закладывается в ранний эмбриональный период (у круп ного рогатого скота на 25—27-е сут) в виде трубчатых выпячи ваний энтодермального покрова в области трех и частично четы рех жаберных карманов первичной глоточной кишки. Затем эти выпячивания превращаются в сплошные тяжи, дающие боковые ответвления — предшественники долек, врастающие в богатую со судами окружающую мезенхиму. Позднее развивающаяся же леза отделяется от жаберных карманов. В конце второго месяца в эпителиальных тяжах появляются лимфоциты, количество ко торых вследствие интенсивного размножения быстро увеличива ется. Постепенно эпителий формирующихся долек приобретает отростчатую форму — создается сеть отростчатых клеток. С треть его месяца в дольках можно различить корковое и мозговое ве щество^ а в последнем появляются первые слоистые эпителиаль ные структуры — тимусные тельца.
Строение. Тимус состоит из своеобразных долек, не являю щихся полностью изолированными образованиями. Совокупность всех долек органа при его реконструкции представляет сложно разветвленные лимфоэпителиальные тяжи, имеющие многочис ленные боковые ответвления. При микроскопии плоскостных срезов с таких ответвлений создается рисунок изолированных долек различной формы и величины, а также долек, соединенных своими основаниями (рис. 206).
Части тимуса покрыты довольно тонкой соединительноткан ной капсулой и широкими междольковыми прослойками, в кото-
282
Рис. 206. Тимус новорожденно го поросенка:
J — капсула; 2 — мозговое ве щество дольки; 3 — корковое ве щество дольки; 4 — менсдольковая соединительная ткань.
рых проходят кровеносные сосуды и содержатся уча стки жировой ткани.
Тимус — лимфоэпителиальный орган. Основу строения долек составляет сеть из отростчатых эпите лиальных клеток — эпителиоретикулоцитов, в про межутках между которыми располагаются и размно жаются многочисленные клетки лимфоидного ряда. В каждой дольке различа
ют |
периферическую часть |
— |
к о р к о в о е в е щ е с т в о и центральную — м о з г о в о е ве |
щ е с т в о , соотношение между которыми в постэмбриональный пе риод изменяется. У новорожденных животных корковое вещество преобладает над мозговым. Большое количество расположенных близко друг к другу ядер лимфоцитов придает корковому веществу характерный вид и темную окраску. Мозговое вещество выглядит более светлым в связи с относительно меньшим количеством лим фоцитов. В этой зоне при световой микроскопии срезов лучше про сматриваются ретикулоэпителиальные клетки (рис. 207). Эпите лиальным клеткам свойственно светлое округлое ядро, содержа щее 2—3 ядрышка и небольшое количество конденсированного хроматина, располагающегося на периферии, около ядерной обо лочки. В цитоплазме имеются мелкие митохондрии, элементы гладкой эндоплазматической сети, комплекс Гольджи; содержат
ся секреторные вакуоли диа метром 0,5—1,5 мкм. Соеди ненные десмосомами эпители альные клетки под капсулой долек и вокруг кровеносных капилляров коркового вещест ва образуют непрерывный
283