

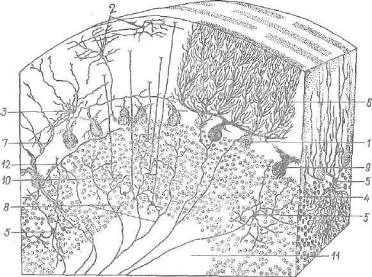
gistologia
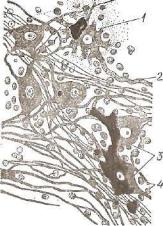
.pdf1—2 — рефлекторные пути сознательных проприоцептивных ощущений и осязания; 8 и 4 — рефлекторные пути проприоцептивных импульсов; б — рефлекторные пути температурной и болевой чувствительности; в — задний собственный пучок; 7 — боковой собственный пучок; 8 — передний собственный пучок; 9 — задний и 10 — передний спинномозжечковые пути; 11 — спинно-таламический путь; 12 — нежный пучок; 13 — клиновидный пучок; 14 — рубро-спинальный путь; 15 — таламоспинальный путь; 16 — вестибуло-спинальный путь; 17 — ретикуло-спинальный путь; 18 — текто-сшгаальный путь; 19 — кортико-спинальный (пирамидный) боко вой путь; 20 — кортико-спинальный пирамидный передний путь; 21 — собственное ядро заднего рога; 22 — грудное ядро (ядро Кларка); 23, 24 — ядра промежуточ ной зоны; 26 — боковое ядро (симпатическое); 26 — ядра переднего рога.
на противоположную сторону спинного мозга в боковой канатик белого вещества, где они образуют вентральный сшганомозжечковый и спино-таламический пути (рис. 176).
Дорсальные рога содержат значительное количество мелких, мультиполярных ассоциативных и комиссуральных нейронов, нейриты которых образуют синапсы на клетках серого вещества спинного мозга той же стороны (ассоциативные клетки) или противоположной (комиссуральные).
Д о р с а л ь н о е |
я д р о образовано крупными клетками, их ак |
|||
соны |
входят в боковой канатик белого вещества той же |
стороны |
||
и в составе |
дорсального спинномозжечкового пути поступают в |
|||
мозжечок. |
|
|
|
|
Нервные |
клетки промежуточной зоны серого вещества обра |
|||
зуют |
два |
ядра: |
м е д и а л ь н о е п р о м е н е у т о ч н о е |
я д р о , |
нейриты которого присоединяются к волокнам вентрального спин
номозжечкового пути той же стороны, и л а т е р а л ь н о е |
про |
м е ж у т о ч н о е я д р о , содержащее ассоциативные клетки |
сим |
патической нервной системы. Аксоны этих клеток через вентраль ные корешки спинного мозга покидают спинной мозг и формируют белые соединительные ветви симпатического ствола.
Ядра вентральных рогов |
серого вещества |
образованы самы |
ми крупными нервными клетками спинного мозга (100—140 мкм |
||
в диаметре). Их нейриты образуют основную |
массу волокон вен |
|
тральных корешков. Через |
смешанные спинномозговые нервы |
|
они поступают в скелетные |
мышцы и заканчиваются в моторных |
|
нервных окончаниях. В вентральных рогах серого вещества спин ного мозга различают две группы моторных клеток: медиальную, иинервпрующую мышцы туловища, и латеральную, которая ха рактерна для области шейного и поясничного утолщений спинлого мозга. Латеральное ядро вентральных рогов содержит нейроциты, иннервирующие мышцы конечностей.
В сером веществе рассеяны нервные клетки, их аксоны в бе лом веществе делятся на более длинную восходящую и более короткую нисходящие ветви. Эти волокна образуют собственные (основные) пучки белого вещества, прилегающие к его серому веществу. Они дают много коллатералей, заканчивающихся си напсами на двигательных клетках передних рогов 4—5 смежных сегментов спинного мозга. В спинном мозге собственных пучков три пары.
Белое вещество спинного мозга состоит из миелиновых нерв ных волокон и опорного нейроглиального остова. Нервные волок на в белом веществе составляют проводящие пути (комплексы волокон) — звенья определенных рефлекторных дуг. Отдельные проводящие пути характеризуются положением и функциональ ной принадлежностью клеток, отростками которых являются их волокна, их синаптическими связями и положением в белом ве ществе спинного мозга.
В числе проводящих путей следует выделить: 1) пути собст венного рефлекторного аппарата спинного мозга, 2) пути, соеди няющие спинной и головной мозг, 3) восходящие (афферент ные) и 4) нисходящие эфферентные (см. анатомию).
ГОЛОВНОЙ МОЗГ
Головной мозг состоит из серого и белого вещества. Серое веще ство располагается на поверхности больших полушарий и обра зует кору большого мозга п мозжечка. Кроме того, оно входит в состав многочисленных ядер ствола мозга.
(1 т и о л м «'л г а включает продолговатый мозг, мост, средний мозг, промежуточный мозг, базальную часть конечного мозга. Ствол имеет внутренний аппарат, обеспечивающий связь между его составными частями. Он содержит ядра, переключающие нервные импульсы, восходящие к коре полушарий и мозжечку и нисходящие от коры на ствол и спинной мозг.
Мультиполяриые нервные клетки ядер серого вещества ство
ла мозга по |
функциональной характеристике |
представлены тре |
|
мя группами: |
моториыад, |
чувствительными |
и ассоциативными. |
М о т о р н ы е |
н е й р о н ы |
аналогичны клеткам вентральных ро- |
|
224 |
15 Заказ №908 |
225 |
|
|
гов спинного мозга. Они формируют двигательные и смешанные ядра черепно-мозговых нервов. Ч у в с т в и т е л ь н ы е я д р а ствола аналогичны нейронам дорсальных рогов спинного мозга. На них оканчиваются аксоны клеток спинальных ганглиев, иду щие в составе дорсального канатика спинного мозга или чувст вительных узлов головы (полулунного, коленчатого, каменистого, яремного и верхнего пучкового). Большое количество ядер обес печивает переключение нервных импульсов из спинного мозга и ствола на кору и от коры на аппарат ствола и спинного мозга (зрительный бугор, зубчатое ядро, оливы и др.).
Продолговатый мозг содержит двигательные и чувствительные ядра собственного аппарата черепных нервов, расположенные преимущественно в области дна IV мозгового желудочка. Двига тельные ядра занимают при этом преимущественно медиальное положение. Чувствительные ядра располагаются по периферии (ядра тонкого и клиновидного пучков, тройничного нерва, слу хового и др.). В продолговатом мозгу локализованы ядра, пере ключающие импульсы на другие отделы головного мозга.
К таким ядрам относятся нижние оливы продолговатого мозга. Они содержат крупные мультиполярные нервные клет ки. Их нейриты образуют синапсы на клетках мозжечка и зри тельного бугра. В нижние оливы поступают волокна от мозжечка» красного ядра, ретикулярной формации и спинного мозга.
Важный координационный аппарат головного мозга — рети* кулярная формация — располагается в центре продолговатого мозга. Здесь в сплетениях нервных волокон лежат мелкие груп пы мультиполярных нейроцитов. Ретикулярная формация рас пространяется по центральной части ствола до промежуточного мозга и является сложным рефлекторным центром.
Белое вещество в продолговатом мозгу располагается преи мущественно вентрально. Основные пучки нервных волокон продолговатого мозга — кортикоспинальные пучки — .пирамиды продолговатого мозга, лежащие в его вентральной части. Спинномозжечковые пути образуют веревчатые тела; поступающие в мозжечок отростки нейронов ядер клиновидного и тонкого пуч ков пересекают серое сетчатое вещество, перекрещиваются по средней линии, образуют шов и направляются к зрительному бугру.
Моет содержит массу поперечно-ориентированных нервных волокон и лежащих между ними нейроцитов.
Средний мозг состоит из серого вещества четверохолмия, по крышки и ножек мозга — массы миелиновых нервных волокон, идущих от коры большого мозга. Покрышка содержит централь ное серое вещество, состоящее из крупных мультиполярных и мелких веретенообразных клеток и волокон.
Промежуточный мозг содержит зрительный бугор. Вентраль но от него располагается гипоталамическая (подбугровая) об ласть. Зрительный бугор включает ядра, отграниченные друг от Друга белым веществом. В вентральных ядрах таламической об-
220
^ШМШШШШяЩШ&ШШШШЩЩЩШШштт.
Рис. 177. Схема строения мозжечка человека:
1 — грушевидные клетки; z — эвездчатые клетки; 8 — корзинчатые клетки; 4 — клетки-зерна; 5 — крупные клетки-зерна (клетки Гольджи); 6 — дендриты гру шевидных клеток; 7 — лазящие волокна; 8 — моховидные волокна; 9 — корзинка полокон; 10 — клубочки мозжечка; и — белое вещество мозжечка; 12 — горизонпип.цые клетки (по Клара).
ласти заканчиваются восходящие чувствительные пути, от них
нервные импульсы передаются коре. |
|
|
|
Гипоталамическая |
область — вегетативный |
центр |
головного |
мозга, регулирующий |
температуру, кровяное |
давление, водный |
|
11 жировой обмен. |
|
|
|
М о к ж о ч о к — орган координации движений и |
равновесия. |
||
Его вффвронтшло и афферентные проводящие пути формируют три пары ножек моожочка. На поверхности органа много изви лин, рмпгрмничоппых глубокими бороздами. Серое вещество об-
раиуот кору мозжечка и ядра, локализованные в белом |
веществе |
||||||||
органа. |
|
|
|
|
|
|
|
|
|
В коре |
мозжечка три слоя, отличающихся но клеточному |
со |
|||||||
ставу: |
наружный — молекулярный, |
средний — ганглиозный |
и |
||||||
нпутрнший — зернистый (рис. 177, 178). |
|
|
|
||||||
Эфферентные |
клетки |
коры |
мозжечка — г а н г л и о з н ы е |
||||||
к л е т к и |
(клетки |
Пуркине) — располагаются в один ряд и |
об |
||||||
разуют ганглиозный слой |
мозжечка. От нерикарионов этих кле |
||||||||
ток в |
молекулярный слой отходят |
2—3 дендрита. |
Последние, |
||||||
разветвляясь в плоскости |
поперечной |
извилине, проходят через |
|||||||
вою толщу молекулярного слоя. От |
противоположного |
полюса |
|||||||
Клеток |
отходят нейриты. Они в виде |
миелиновых волокон в со |
|||||||
ставе |
белого вещества следуют к |
ядрам мозжечка. |
В |
пределах |
|||||
15* |
|
|
|
|
|
|
|
|
227 |
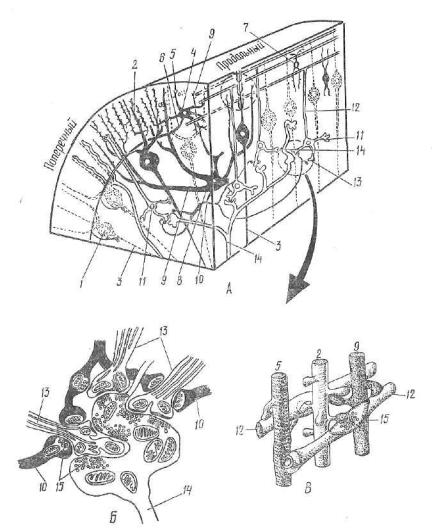
А — извилины мозжечка (пунктиром на продольном срезе ограничены участки, показанные на рисунках Б и В); Б — клубочек зернистого слоя; В — синапсы в молекулярном слое; 1 — грушевидная клетка (клетка Пуркине); 2 — дендриты грушевидных клеток; з — нейрит грушевидной клетки; 4 — корзинчатые клетки; 5 — дендриты и б — аксон корзинчатой клетки; 7 — звездчатая клетка; 8 — крупные клетки-зерна (клетки Гольджи); 9 — дендриты клетки Гольджи; ю — аксон клетки Гольджи; 11 — клетки-зерна; 12 — аксон клетки-зерна; 13— дендриты клеток-зерен; 14 — моховидные волокна; 15 — синаптические пузырь ки. В черный цвет окрашены терминальные нейроны (по Сантогатай).
зернистого слоя нейриты отдают коллатерали, которые, возвра щаясь в ганглиозный слой, вступают в синаптическую связь с грушевидными клетками.
Молекулярный слой коры мозжечка содержит корзинчатые и |
|
:шездчатые тормозные |
нейроны. К о р з и н ч а т ы е к л е т к и ле |
жат в глубокой зоне |
молекулярного слоя. Они имеют небольшие |
размеры перикарионов (10—20 мкм), тонкие длинные дендриты |
|
и аксоны, ориентированные поперек извилин, следующие на боль
шие расстояния |
над телами грушевидных клеток и образующие |
|||||
с ними |
синаптические |
связи — «корзинки», оплетающие перика- |
||||
рион. |
Многочисленные |
з в е з д ч а т ы е |
к л е т к и |
молекулярного |
||
слоя коры |
мозжечка |
представлены двумя видами: мелкими и |
||||
крупными клетками. Мелкие звездчатые клетки |
имеют короткие |
|||||
дендриты. |
Их нейриты |
образуют синапсы на дендритах груше- |
||||
видных |
клеток. |
Крупные звездчатые |
клетки |
характеризуются |
||
длинными сильно разветвленными нейритами и дендритами. Тер минальные ветви их дендритов контактируют как с дендритами, так и с телами грушевидных клеток, включаясь в состав их перицеллюлярных «корзинок». Звездчатые и корзинчатые клетки передают тормозные импульсы на грушевидные клетки коры моз жечка в плоскости поперечной извилине.
В зернистом слое коры мозжечка различают клетки-зерна и два вида клеток Гольджи (с короткими и длинными нейритами). IV л етки-з е р н а имеют маленький по объему, бедный цитоплаз мой: перикарион с круглым крупным ядром. Характерные для них 3—4 коротких дендрита заканчиваются ветвлением в виде лапки птицы. Последние образуют синапсы с окончаниями при ходящих в кору мозжечка возбуждающих моховидных волокон. Нейриты клеток-зерен поступают в молекулярный слой коры и в нем Т-образно делятся на две ветви, ориентированные вдоль извилин органа, параллельно его поверхности. Имея большую протяженность, они пересекают зоны ветвления многих рядов грушевидных клеток и соответственно образуют многочисленные синапсы как с ними, так и с дендритами корзинчатых и груше видных клеток молекулярного слоя. Нейриты клеток-зерен проподят возбуждение, полученное ими от моховидных волокон, и передают ого по параллельным волокнам (ветвлениям своих ден дритов) на значительное расстояние грушевидным клеткам с по мощью синапсов на их дендритах.
З в е в д ч а т ы е к л е т к и Г о л ь д ж и представлены двумя видами — звездчатые клетки с короткими и длинными нейритами. С короткими отростками клетки лежат вблизи ганглиозного слоя. Их дендриты в молекулярном слое образуют синапсы с парал
лельными волокнами (с аксонами клеток-зерен). |
Нейриты |
идут |
в зернистом слое и образуют тормозные синапсы |
на концевых |
|
отделах дендритов клеток-зерен проксимальнее |
синапсов |
мохо |
видных волокон, |
блокируя поступление возбуждающих импуль |
||
сов с моховидных |
волокон на клетки-зерна |
(7), Клетки |
Гольджи |
с длинными отростками характеризуются |
ветвящимися |
в зерни- |
|
229
втом слое |
дендритами и |
нейритом, уходящим |
в белое |
вещество |
||
мозжечка. |
|
|
|
|
|
|
В кору |
мозжечка |
поступает два вида афферентных волокон — |
||||
моховидные |
и лазающие. |
|
|
|
|
|
М о х о в и д н ы е |
в о л о к н а , |
поступая из |
белого |
вещества |
||
мозжечка в |
кору, заканчиваются |
синапсами в |
«клубочках» моз |
|||
жечка, контактируя |
с |
терминальными ветвлениями |
дендритов |
|||
клеток-зерен (14). Нейриты последпих по параллельным волок нам проводят импульс в молекулярпый слой коры к дендритам грушевидных, корзинчатых, звездчатых клеток и клеток Гольджи.
Л а з а ю щ и е в о л о к н а , пересекая зернистый слой, подхо дят к грушевидным клеткам и по их дендритам поднимаются в молекулярный слой, заканчиваясь на них синапсами.
Следовательно, возбуждающие импульсы в кору мозжечка по ступают на грушевидные клетки или по лазающим волокнам не посредственно, или по моховидным волокнам через клетки-зерна яо параллельным волокнам (их аксоном).
Торможение эфферентных нейронов коры (грушевидных клеток) обеспечивается возбуждением звездчатых, корзинчатых клеток и клеток Гольджи. Поступление в кору мозжечка воз буждения по моховидным волокнам через клетки-зерна может быть прервано тормозными синапсами клеток Гольджи, локали зованными на терминальных ветвлениях дендритов клеток-зерен проксимальнее их возбуждающих синапсов с моховидными во локнами.
Глиальные элементы коры мозжечка представлены волокни стыми и плазматическими астроцитами. В ее ганглиозном слое между грушевидными нейроцитами лея^ат клетки глии, отли
чающиеся |
темными ядрами. Их отростки |
образуют |
«бергма - |
н е в с к и е |
в о л о к н а » молекулярного слоя |
мозжечка, |
располо |
женные |
между ветвями дендритов ганглиозпых клеток. В моле |
|||
кулярном |
и ганглиозном |
слоях коры |
мозжечка |
сосредоточены |
г л и а л ь н ы е м а к р о ф а г и . |
|
|
||
К о р а |
п о л у ш а р и й |
г о л о в н о г о |
м о з г а |
образована се |
рым веществом, состоящим из нескольких слоев клеток, различ ных по форме, размерам и функциональному значению. Наибо лее специфичны для коры полушарий нервные клетки пирами дальной формы. Они характеризуются вытянутой треугольной формой перикарионов с вершиной, обращенной к поверхности мозга. От расширенного основания тела клетки, обращенного к белому веществу мозга, отходит нейрит, заканчивающийся сирапсом в сером веществе смежного участка мозга или выходя щий в белое вещество, формируя проводящие пути центральной нервной системы.
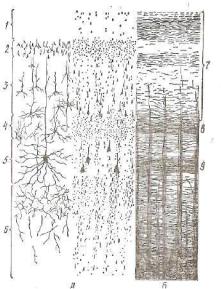
Близкие по форме, размерам и функциональному значению нервные клетки образуют нерезко разграниченные слои коры, характерные для различных отделов полушарий головного моз га. В двигательной эоне полушарий различают шесть основных слоев: молекулярный, наружный зернистый, пирамидный, внут-
230
Рис. 179. Кора больших полуша рий мозга:
Л — схема расположения клеток (цитоархитектоника); В — схема распо ложения волокон (миелоархитектоника в наиболее развитых отделах коры); J — молекулярный слой; 2— наружный зернистый слой; 3 — пи рамидный слой; 4 — внутренний зер нистый слой; б — ганглионарный слой; 6 — слой полиморфных кле ток; 7 — наружный главный слой подокон; & — наружная полоска Байарже; 9 — внутренняя главная полос ка (по Баргману).
рсиний зернистый, ганглиозный, слой полиморфных кле ток (рис. 179).
Молекулярный слой содер жит сеть нервных волокон и отдельные мелкие нервные клетки, преимущественно ве ретенообразной формы. Нерв ные волокна ориентированы
параллельно поверхности мозга. Большая часть их представляв* собой отростки нижележащего слоя.
В наружном зернистом слое — мелкие нервные клетки диамет ром около 10 мкм, округлой, звездчатой или пирамидальной фор мы. Их дендриты направляются в молекулярный слой. Нейриты частично уходят в белое вещество, частично также поступают в сплетение волокон молекулярного слоя.
Пирамидный слой особенно хорошо развит в прецентралыюй извилине. Клетки этого слоя имеют пирамидальную форму. Разморы их последовательно увеличиваются от 10 мкм в его пери ферической зоне до 40 мкм в центральной. От верхушки клетки отходит дендрит, конечные ветвления которого располагаются в молекулярном слое. Дендриты боковой поверхности и основания клотки короткие и образуют синапсы со смежными клетками Втого слоя. Нейрит отходит от основания клетки. Нейриты более мелких клеток не выходят эа продолы коры, а у более крупных пирамид в виде миелиновых, ассоциативных или комиссуральных волокон уходят в белоо вещество.
Внутренний зернистый слой в различных полях коры выра« жен по-ра8иому. В зрительной зоне коры он сильно развит, а в ее двигательной зоне слабо. Внутренний зернистый слой образо ван мелкими звездчатыми и пирамидальными клетками. В нем много горизонтальных волокон.
Ганглиозвый слой коры содержит крупные клетки пирами* дальной формы до 120 мкм высоты и 80 мкм ширины в основа* нии пирамиды. В цитоплазме клеток наблюдают крупные базофильные глыбки, что свидетельствует о высоком уровне синтеза
231
белков, необходимых для поддержания массы цитоплазмы их длинных аксонов, формирующих кортикосшшальные пути спин ного мозга.
В слое полиморфных клеток нейроны различной формы и раз меров. Клетки в большем количестве и более крупные сосредо точены во внешней зоне этого слоя. Отростки клеток длинные. Их нейриты уходят в белое вещество в составе эфферентных пу тей головного мозга. Дендриты достигают молекулярного слоя коры.
ВЕГЕТАТИВНЫЙ ОТДЕЛ НЕРВНОЙ СИСТЕМЫ |
|
|
||||||
Вегетативный отдел |
нервной системы |
включает высшие |
вегета |
|||||
тивные центры, локализованные в промежуточном мозге |
в обла |
|||||||
сти |
III желудочка, |
вегетативные |
ядра |
серого |
вещества |
ствола |
||
и спинного мозга, а также периферические |
нервные ганглии. |
|||||||
|
В отличие от рефлекторных дуг соматической нервной систе |
|||||||
мы моторный нейрон рефлекторных дуг |
вегетативного |
отдела |
||||||
идет не в составе |
серого вещества |
центральной нервной системы, |
||||||
а в |
одном из периферических ганглиев. |
|
|
|
|
|||
|
Вегетативная |
нервная система |
представлена |
парасимпатиче |
||||
ской и симпатической системами. |
К парасимпатической нервной |
|||||||
системе относят нейроны черепно-мозгового и крестцового уча стков мозга и связанные с ними ганглии. Симпатическая нервная система объединяет нейроны грудопоясничного отдела мозга и связанные с ними превертебральные и паравертебральные ган глии. Симпатические нервные волокна иниервируют все органы, тогда как парасимпатическая нервная система иннервирует лишь органы, развившиеся из эмбриональной кишки или в связи с ней.
Большинство внутренних органов получает как симпатиче ские, так и парасимпатические нервные волокна.
Эфферентное звено вегетативных рефлекторных дуг двух членно. Центральный нейрон всегда располагается в головном
или спинном мозге. |
Второй нейрон |
(периферический) |
в симпа |
|||
тической нервной системе |
лежит в |
превертебральных или пара- |
||||
вертебральных узлах, |
а в |
парасимпатической — в органе |
или |
|||
вблизи него (рис. 180). |
|
|
|
|
||
В |
п а р а с и м п а т и ч е с к о й н е р в н о й с и с т е м е |
оба ней |
||||
рона |
холинергические. Пресинаптический полюс как первого, так |
|||||
и второго нейрона содержит мелкие, |
светлые синаптические |
пу |
||||
зырьки (40—60 нм) и одиночные крупные электроноплотные ве зикулы.
В симпатической нервной системе первый нейрон холинергический, второй адренергический (медиатор — иорадреналин). Морфологически адренергические синаптические пузырьки (30— 50 нм в диаметре) характеризуются электроноплотной сердцевиной (гранулярные синаптические пузырьки). Кроме характерных для адренергических синапсов гранулярных везикул, в их составе, как и в холинергических, выявляют и большие гранулярные ве-
Гис.
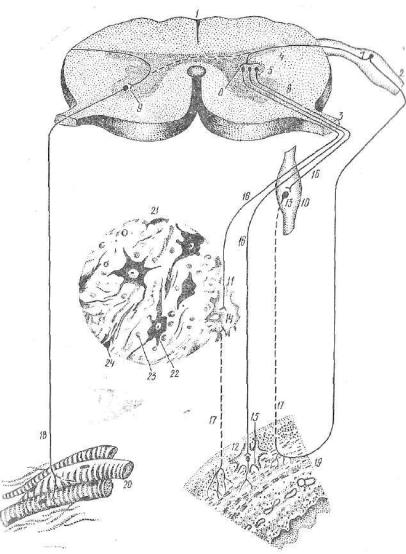
/ — спинной мозг: 2 — спинномозговой узел; 8 — передний корешок; 4 — задний
.„„.• s _ боковой рог; 6 — передний рог; 7 — чувствительный (афферентный) ней- \чш соматической и симпатической нервной системы; 8 — центральный (эфферент- П.1Й) нейсон вегетативной нервной системы; 0 — двигательный (эфферентный) нейiioii пеоешщх рогов- Ю — узел симпатического ствола; 11 — узел солнечного спле тения- 18 —"интрамуральный узел (узел нервного сплетения пищевода); 13 14, 15— периферические (эфферентные) нейроны вегетативной нервной системы; 16 — пре ти г. пленарные волокна эфферентного пути вегетативной нервной системы; 17 — моотганглионарные волокна эфферентного пути вегетативной нервной системы, is— иффепентный путь соматической нервной системы; 19 — стенка пищевода; го — поти.очнополосатыв мышцы; 21 — микроскопическое строение периферических узлов ипгетативной нервной системы; 22 — мультиполярная нервная клетка; 23 — клетки Ший; 24 — нервное волокно.
232
|
|
Рис. |
181. |
Нервные |
клетки вегетативного |
|||||
|
|
ганглия: |
|
|
|
|
|
|
|
|
|
|
1 — длинноаксонная нервная клетка; |
2 — ак |
|||||||
|
|
сон; з — равноотростчатая нервная клетка; 4 — |
||||||||
|
|
ядра клеток глии. |
|
|
|
|
|
|
||
|
зикулы диаметром |
60—120 |
нм |
(1— |
||||||
|
5 % к общему количеству). |
|
|
|
||||||
|
|
В с и м п а т и ч е с к и х н е р в н ы х |
||||||||
|
узлах, кроме свойственных для них |
|||||||||
|
мультиполярных нейронов, |
имеются |
||||||||
|
группы |
мелких |
гранулосодержащих |
|||||||
|
интенсивно флуоресцирующих клеток |
|||||||||
|
|
(МИФ-клетки). Для них типичны ко |
||||||||
|
роткие отростки |
и |
наличие |
в |
со |
|||||
|
ставе |
цитоплазмы гранулярных |
пу- |
|||||||
|
вырьков. |
По флуоресценции и суб |
||||||||
микроскопическому строению |
их |
пузырьки |
соответствуют |
пу |
||||||
зырькам цитоплазмы |
клеток |
мозгового вещества |
надпочечни |
|||||||
ков. Предполагается |
их участие в проведении |
нервного |
|
им |
||||||
пульса от преганглионарных |
волокон на нейроны |
ганглия |
(роль |
|||||||
интернейронов). МИФ-клетки рассматривают как внутриганглионарную тормозную систему. Возбуждаясь преганглионарными синапсами, они выделяют катехоламины, тормозящие передачу
нервного импульса с преганглионарных |
волокон на симпатиче |
|
ские нейроны узла. |
|
|
Й а т р а м у р а л ь н ы е н е р в н ы е |
с п л е т е н и я . |
Значи |
тельное количество нейроцитов вегетативной нервной |
системы |
|
сосредоточено в нервных сплетениях полых органов: пищевари тельного тракта, сердца, мочевого пузыря ж др. Нервные узлы сплетений содержат эфферентные, реценторные и ассоциативные
нейроны. |
Морфологически |
в |
интрамуральных нервных |
узлах |
||||
различают три типа нервных клеток |
(рис. 181). |
|
|
|||||
К л е т к и |
п е р в о г о |
т и п а |
Д о г е л я (длинноаксонпые нев- |
|||||
роциты) |
характеризуются длинным аксоном и многими ветвящи |
|||||||
мися депдритами. К л е т к и |
в т о р о г о т и п а |
Д о г е л я (равно- |
||||||
отростчатые |
нейроциты) |
содержат |
несколько |
отростков, |
среди |
|||
которых морфологически нельзя определить аксон. Эксперимен
тально установлено, что он заканчивается |
синапсом на клетках |
||
первого типа. К л е т к и |
т р е т ь е г о т и п а |
образуют |
синаптиче- |
ские связи с депдритами |
нейронов соседних ганглиев. В стенке |
||
желудочно-кишечного тракта имеется три |
нервных |
сплетения: |
|
подсерозное, межмышечное и подслизистое, содержащие ганглии нервных клеток.
Наиболее массивное межмышечное нервное сплетение распо ложено в мышечной оболочке органа между продольным и цир кулярным слоями. Нейрогистологически, электронно-микроскопи- чески, гистохимически и нейрофизиологически выявлена специ фичность этого сплетения» позволяющая сопоставить его по ряду
признаков с центральной нервной системой. В частности, меж^ мышечное нервное сплетение кишечника также покрыто соеди*> пительнотканной оболочкой, отграниченной от нервной ткани ба* зальной мембраной. Нервное сплетение имеет свою систему кровоснабжения в виде собственной капиллярной сети, локализо ванной за пределами капсулы. Капилляры и соединительная ткань в паренхиму ганглия не проникают.
К а п с у л а с п л е т е н и я содержит 2—3 слоя коллагеновых волокон, разграниченных друг от друга плоскими клетками. Внут* ри каждого слоя волокна ориентированы параллельно, не обра* зуя пучков. В цитоплазме клеток капсулы различают свободные рибосомы, митохондрии и пиноцитозные пузырьки. Последние свидетельствуют об участии этих клеток в процессах транспорта веществ.
Экспериментами показано, что эндотелий капилляров п кап сула сплетений участвуют в формировании барьера «кровь —•- нервное сплетение», препятствующего проникновению в послед-» нее молекул маркеров.
Н е й р о г л и я г а н г л и е в межмышечного нервного сплетв* иия не дифференцирована (в отличие от экстрамуральных узлов)] на сателлиты капсулы нейроцитов и леммоциты волокон. Глио* циты одновременно граничат с перикарионами нейронов, покры* вают группы осевых цилиндров и синаптические образования» Цитоплазма глиоцитов бедна органеллами. Они содержат неболь* шие цистерны гранулярной эндоплазматической сети, одиночные митохондрии и свободные полирибосомы.
Н е й р о ц и т ы г а н г л и е в |
межмышечного нервного сплете* |
пия характеризуются обилием |
органелл. Крупные комплексы |
Гольджи и цистерны эндоплазматической сети в совокупности образуют в клетках плотную сеть мембран. Многочисленные ри* босомы локализированы как свободно, так и па мембранах эндо-- плазматической сети. Гранулярпая эндоплазматическая сеть рас* иределена равномерно и не образует хроматофильных глыбок.
В ганглиях межмышечного нервного сплетения кишечнике содержатся эфферентные и афферентные нейроны (рис. 182). Ме тодом флуоресценции и электронной микроскопии на клетках первого типа Догеля выявляются возбуждающие холипергиче-3 ские и тормозные адренергические синапсы.
За небольшим исключением, иптрамуральные нейроны кшпеч* пика неадренергические, но они снабжены адренергическими си* папсами главпым образом аксонов нервных клеток преверте* бральных ганглиев. Электронно-микроскопически эти синапсы характеризуются типичными для адреиергических элементов гра нулярными синаптическими пузырьками (30—60 нм).
Методом флуоресцентной микроскопии установлено, что адрепергические синапсы имеют большинство нейронов сплетения,. Отсутствие флуоресцирующих волокон при перерезке брыжееч* пых нервов свидетельствует, что адренергические синапсы в со ставе интрамуральных сплетений образованы окончаниями аксо-
234 |
23! |
|
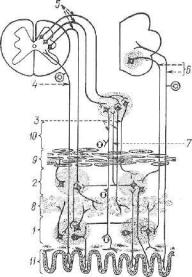
Рис. 182. Схема интрамуральных сплетений пищеварительного тракта: 1 — подслизистое сплетение; г — миэнтеральное сплетение; з — постганглионарные симпатические волокна; 4 — спинальные афферентные волокна; 5 — симпатические преганглионарные волок на; 6 — вагусные афферентные волокна; 7 — афферентные волокна местных реф лекторных путей; 8 — кольцевой мышеч ный слой; 9 — продольный мышечный слой; 10 — серозный слой; 11 — слизи стая оболочка (по Ноздрачеву, 1978).
нов нейроцитов преили паравертебральных ганглиев.
В кишечнике, кроме холинергических нейроцитов, имеют ся тормозящие пуринергические нейроны, выделяющие в качест ве медиатора пуриновые соеди нения. Перикарионы и их отро стки содержат характерные для них электроношготные пузырьки
100 нм в диаметре. Пуринергические нейроны вызывают нисхо дящее торможение пищеварительного тракта, что является звеном перистальтического рефлекса. В отличие от пуринергических адренергические нервы вызывают рефлекторное угнетение пери стальтики торможением интрамуральных холинергических воз буждающих нейронов (Береток и Коста, 1979).
Центральная нервная система получает информацию о внешнем
мире |
и |
о внутреннем |
состоянии |
организма от специализирован |
||
ных |
к |
восприятию |
раздражений |
разнообразных |
рецепторыых |
|
приборов. В соответствии с |
местом расположения |
различают |
||||
интеро- и экстерорецепторы. |
И н т е р о р е ц е п т о р ы |
служат для |
||||
восприятия специфических раздражений из внутренней среды
организма: висцерорецепторы сигнализируют о состоянии внут |
|
ренних органов; проприорецепторы |
воспринимают раздражения |
от органов произвольного движения |
(кости, мышцы, связки, су |
ставы) ; вестибулорецепторы посылают импульсы о положении
тела или его отдельных |
частей в пространстве. |
Э к с т е р о р е |
|
ц е п т о р ы |
воспринимают раздражения, поступающие из внеш |
||
ней среды. |
К ним относят зрительные, слуховые, |
обонятельные, |
|
вкусовые и осязательные |
рецепторы. |
|
|
Рецептор — только часть единой системы, которую И. П. Пав лов обозначил термином а н а л и з а т о р . Каждый анализатор имеет три отдела: периферический (воспринимающий), средний
236
(проводящий) и центральный (определенный отдел коры полу шарий головного мозга, куда поступает информация и где про исходит ее декодирование и последующее использование при фор мировании многочисленных ответных реакций организма).
Органы чувств — это периферические части анализаторов, со
держащие собственно рецепторные элементы, в |
которых проис |
|||
ходит преобразование |
сигналов |
внешнего мира |
в |
нервный им |
пульс. Вместе с разнообразными |
вспомогательными структурны |
|||
ми образованиями они |
морфологически оформлены |
как сложно |
||
устроенные специализированные |
органы — органы |
зрения, слуха |
||
и др. |
|
|
|
|
Содержащиеся в органах чувств рецепторные |
клетки снабже |
|||
ны особыми подвижными универсальными антеннами, автомати чески воспринимающими те или другие виды внешней энергии (жгутиками — киноцилиями или микроворсинками — стереоцилиями). Пусковым механизмом для общего возбуждения рецепторной клетки является первичное, специфическое взаимодейст вие энергетической единицы стимула с особой белковой молеку лой на биологической мембране этой антенны. Такое взаимодей ствие по своей чувствительности нередко приближается к физическому пределу различения данного стимула. Например, палочки в сетчатке глаза могут воспринимать единичный фотон света.
В соответствии с особенностями развития и строения в пери ферическом отделе анализатора воспринимающими элементами
могут |
быть клетки |
двух |
разновидностей: |
п е р в и ч н о ч у в с т - |
||||||
в у ю щ и е — нервные клетки |
и |
в т о р и ч н о ч у в с т в у ю щ и е — |
||||||||
оеисоэпителиальные |
клетки. Первичночувствующие |
клетки |
пре |
|||||||
образуют сигналы из внешпей |
среды с помощью |
высокоспециали- |
||||||||
-зированного периферического |
отростка — видоизмененного |
ден |
||||||||
дрита. |
С помощью |
центрального |
отростка |
( а к с о н а ) |
возбужде |
|||||
ние передается на |
последующие |
нервные |
клетки |
анализатора. |
||||||
К органам, имеющим |
первичночувствующие |
клетки, |
относят |
|||||||
орган |
зрения и орган обоняния. |
|
|
|
|
|
|
|||
Рецепторными элементами в органах слуха, вкуса и равнове сия являются вторичночувствующие клетки. С участием особого питейного аппарата возбуждение, возникшее в этих клетках, нородается иа концевые участки деидритов чувствительных нервпых клеток, расположенных в специальных ганглиях, и далее в другие нервные клетки промежуточного отдела анализатора.
В данной главе рассмотрены органы зрения, слуха и равно весия. Рецепторы обоняния, вкуса и осязания изложены вместе с органами, в которых они заключены.
ОРГАН ЗРЕНИЯ
Орган зрения — глаз представляет собой периферическую часть зрительного анализатора. Состоит он из глазного яблока, соеди ненного посредством зрительного нерва с головным мозгом, за-
237
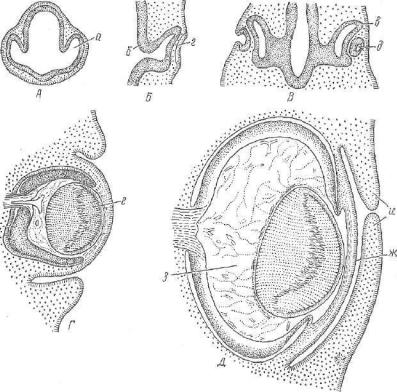
Рис. 183. Схема развития глаза:
А, В, В, Г, Д — раэрезы эмбрионального зачатка глаза кролика на разных стади ях развития (по Бремеру); о — глазной пузырь; б — глазной стебелек; в — глаз ной бокал; в — хрусталиковая пластинка; 9 — хрусталиковый пузырек; е — хруста* лик; ж — роговица; а — стекловидное тело; и — веки.
щитных и вспомогательных органов — веки, слезный аппарат, поперечнополосатые глазодвигательные мышцы, фасции.
Развитие. Различные структурные элементы глазного яблока развиваются из разных эмбриональных зачатков. В начале воз никает выпячивание стеики промежуточного мозга — глазной пузырь, растущий по направлению к эктодерме и соприкасаю щийся с нею. В месте соприкосновения эктодерма образует утол щение — хрусталиковую пластинку. Та часть глазного пузыря, которая оказывается в контакте с хрусталиковой пластинкой, впячивается, в результате чего глазной пузырь превращается в двухслойный глазной бокал. Край глазного бокала становится аачатком радужной оболочки и цилиарного тела, наружный слой остальной части — зачатком пигментного эпителия сетчатки, а
внутренний слой — ее светочувствительной частью (рис. 183).
Одновременно с развитием глазного яблока хрусталиковая пластинка сама впячивается в полость глазного бокала в виде мешочка, который отсоединяется от эпителия и превращается в хрусталиковый пузырек. Клетки его, обращенные к сетчатке, сильно вытягиваются и превращаются в первичные хрусталиковые волокна, заполняющие полость пузырька. Эпителий, распо ложенный над хрусталиком, перерастает в эпителий роговицы.
Наружные оболочки глаза (сосудистая и склера) образуются из мезенхимы, окружающей развивающийся глазной бокал. Кле точные элементы миотомов области головы участвуют в форми ровании мышц глазного яблока. Таким образом, при развитии глаза многие части, из которых возникает глазной бокал, всту пают между собой в сложные индукционные взаимоотношения.
ГЛАЗНОЕ ЯБЛОКО
Стенка глазного яблока состоит из трех оболочек — наружной, средней и внутренней. В составе глазного яблока имеются также светопреломляющие образования и среды — хрусталик, жидкость передней и задней камер глаза, стекловидное тело (рис. 184).
Н а р у ж н а я оболочка |
имеет две |
части — роговицу и |
склеру. |
|
|
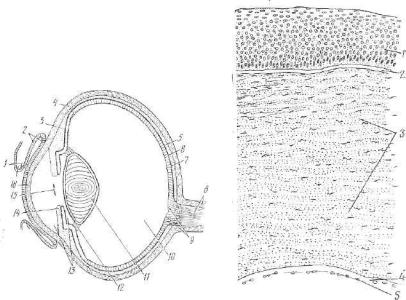
Роговица (сотпеа) — передняя часть наружной оболочки, со |
||
стоит из переднего эпителия |
роговицы, |
базальной мембраны, |
передней пограничной мембраны, собственного вещества рого
вицы, задней пограничной мембраны и заднего эпителия |
рого |
вицы (рис. 185). |
|
Многослойный плоский неороговевающий п е р е д н и й |
эпи |
т е л и й (epithelium anterins) состоит из 5—7 слоев клеток. В нем содержатся многочисленные рецепторные окончания, придающие роговице большую тактильную чувствительность (рефлекс рого вицы). Вазальные клетки эпителия обладают выраженной митотической активностью, поэтому при повреждении эпителий ро
говицы |
быстро восстанавливается. |
Передний эпителий |
продол |
||
жается |
в эпителии конъюнктивы |
и увлажняется |
секретом |
||
слезных и конъюнктивальных желез. |
|
|
|||
Б а з а л ь н а я |
м е м б р а н а — гомогенная |
белково-полисаха- |
|||
ридная пластинка. |
|
|
|
|
|
П е р е д н я я п о г р а н и ч н а я |
м е м б р а н а (боуменова) у |
||||
разных животных |
имеет различную толщину; |
особенно выраже |
|||
на в роговице крупного рогатого скота. Электропно-микроскопи-
чоски в составе этой мембраны |
обнаружены тонкие коллагено- |
иые фибриллы, войлокообразно переплетающиеся. |
|
С о б с т в е н н о е вещество |
роговицы (substantia pro |
pria согнеае) — основная масса роговицы. Состоит из многочис ленных, правильно чередующихся соединительнотканных пласти нок, каждая из которых имеет параллельно расположенные пуч ки коллагеновых фибрилл. Между пластинками залегают упло щенные клетки фибробластического ряда и содержится аморф-
238
239
ный компонент. Прозрачности собственного вещества способствует то, что все его коллагеновые фибриллы параллельно ориенти
рованы |
и одинаковы по толщине, благодаря чему |
свет, который |
|||||||||
они рассеивают, гасится в результате |
интерференции. Этому же |
||||||||||
способствуют |
сульфатированные гликозаминогликаны, |
которые |
|||||||||
обусловливают |
способность ткани к набуханию |
и |
поддерживают |
||||||||
упорядоченное |
|
расположение |
фибрилл. Место |
перехода |
собст |
||||||
венного |
вещества роговицы в |
соединительную |
|
ткань |
склеры |
на |
|||||
зывается |
л и м б |
(limbus — край). В |
области |
лимба |
находится и |
||||||
край передней |
пограничной мембраны, то есть она не переходит |
||||||||||
с роговицы па |
склеру. |
|
|
|
|
|
|
|
|
||
З а д н я я |
п о г р а н и ч н а я |
м е м б р а н а |
(десцеметова) |
све- |
|||||||
томикроскопически представляет гомогенную пластинку. Электрон- но-микроскопически в ней обнаружены коллагеновые фибриллы, которые, пересекаясь, образуют шестиугольные фигуры.
П л о с к и й э п и т е л и й з а д н е й п о в е р х н о с т и р о г о в и ц ы состоит из одного слоя клеток, имеющих шестигранные и другие очертания. Этот эпителий переходит в эпителий, покрываю щий переднюю поверхность ра дужной оболочки.
Роговица обеспечивает значи тельный процент фокусирующей
Рис. 184. Схема строения глазного яб |
Рис. 185. Роговица глаза телеп- |
||
лока: |
|
|
ка: |
1 — край верхнего века с ресницей; 2 — |
1 — многослойный ПЛОСКИЙ эпи |
||
конъюнктива; з — ресничное тело и рес |
телий; 2 — передняя пограничная |
||
ничная часть сетчатки; 4 — склера (белоч |
мембрана; 3 — собственное ве |
||
ная оболочка); |
5 — сосудистая оболочка; |
щество роговицы; 4 — задняя пос |
|
в — сетчатка; 7 — пигментный слой; 8 — |
траничная мембтэана; 5 — одно |
||
зрительный нерв и его оболочки; 9 — сосо |
слойный плоский эпителий. |
||
чек зрительного |
нерва; 10 |
— стекловидное |
|
тело; 11 — хрусталик; 12 — хрусталико- |
|
||
вая связка; 13 — передняя и задняя каме |
|
||
ры глаза; 14 — радужная |
оболочка; 15 — |
|
|
зрачок; 16 — роговица. |
|
|
|
240 |
|
|
|
способности глаза и действует, как сильная лупа. Это особая} часть глаза, у которой нет кровеносных капилляров, питание она получает из передней камеры глаза и сосудов лимба. Таким об разом, роговица достаточно изолирована и именно благодаря этому обстоятельству возможна ее пересадка от одного организма, другому.
В случае повреждения роговицы и воспалительного процесса происходят врастание в нее кровеносных капилляров, проникно вение клеток (лейкоцитов и др.), что вызывает нарушение опти ческих свойств и помутнение роговицы.
Склера (sclera — твердый) — непрозрачная задняя и переднебоковая части наружной оболочки, белого цвета, самая прочная i; стенке глазного яблока. Состоит из плотной соединительной ткани, в которой коллагеновые волокна и образованные из них пластинки расположены параллельно поверхности глаза. Между
ними |
находятся |
эластические волокна и уплощенные фибробла- |
|
сты. |
|
|
|
На |
границе с |
роговицей |
образуется утолщение склеры, хо |
рошо |
видимое в виде в а л и к а у плотоядных. В ткани склеры за |
||
валиком имеются |
небольшие |
разветвленные полости — в е н о з |
|
ное с п л е т е н и е , обеспечивающее отток жидкости из передней
камеры глаза. |
В задней части |
склеры |
имеется р е ш е т ч а т а я |
п л а с т и н к а |
(lamina cribrosa), |
содержащая мелкие отверстия,, |
|
через которые |
проходят нервные волокна, формирующие зритель |
||
ный нерв. Снаружи склера покрыта |
эписклеральной рыхлой |
||
соединительной тканью, содержащей многочисленные капил ляры.
|
Склера выполняет функцию прочного остова стенки глаза, в |
||||||||||||
нее |
проникают, а затем прикрепляются к ней волокна сухожилий |
||||||||||||
глазных |
мышц. |
|
|
|
|
|
|
|
|
|
|||
|
У птиц в связи с неполным |
окостенением стенок орбиты скле |
|||||||||||
ра |
около роговицы |
содержит |
черепицеобразио |
расположенные |
|||||||||
мелкие |
костные чешуйки, которые, соединяясь, |
образуют своеоб |
|||||||||||
разное защитное кольцо. Дистально от |
него |
до |
места |
впадения |
|||||||||
зрительного |
нерва |
в |
склере |
находится |
гиалиновая |
хрящевая |
|||||||
ткань. |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
С р е д н я я |
о б о л о ч к а состоит |
из |
трех |
частей: |
радужной |
|||||||
оболочки, ресничного тела и сосудистой |
оболочки |
(рис. 186). |
|||||||||||
|
Радужная |
оболочка |
(iris) — передняя часть средней оболочки. |
||||||||||
Пространство |
между |
радужной оболочкой |
и |
роговицей |
называ |
||||||||
ется п е р е д н е й к а м е р о й |
г л а з а , |
а |
между |
радужной обо |
|||||||||
лочкой |
и |
хрусталиком — з а д н е й |
к а м е р о й . |
В центральной |
|||||||||
части радужной оболочки имеется отверстие — з р а ч о к , |
который |
||||||||||||
у собак, свиней и птиц |
округлой формы, у кошки в виде верти |
||||||||||||
кальной щели, у травоядных поперечно-овальный. Задний край радужной оболочки, соединяющий ее с ресничным телом, назы вают р е с н и ч н ы м к р а е м . Основу радужной оболочки состав ляют пучки клеток гладкой мышечной ткани и рыхлая соедини
тельная ткань с большим |
количеством пигментных клеток — хро- |
16 Заказ Ш 908 |
241 |
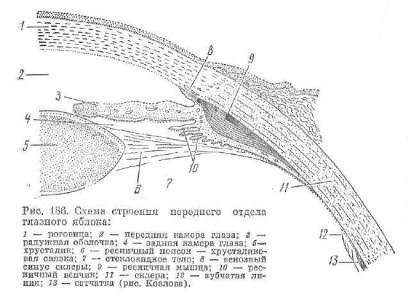
матофоров и многочисленными кровеносными сосудами. На •поперечном разрезе радужной оболочки по направлению от пе редней к задней поверхности различают пять слоев: эпителиаль ный, наружный пограничный, сосудистый, внутренний погранич ный и пигментный. Последний является продолжением пигмент ного эпителия ресничного тела и далее сетчатки. Во всех слоях радужной оболочки в разном количестве имеются пигментные клетки, которые обусловливают цвет глаз. У животных альби носов пигментных клеток нет, поэтому у них радужная оболочка красного цвета, в связи с тем что через ее толщу просвечивают кровеносные сосуды.
Гладкая мышечная ткань формирует в радужной оболочке две мышцы. Мышца, суживающая зрачок ( с ф и н к т е р ) , состоит из пучков клеток, ориентированных циркулярно и расположенных вблизи зрачкового края оболочки. Пучки клеток, расширяющих
зрачок ( д и л я т а т о р ) , |
имеют |
радиальное |
направление и нахо |
|||
дятся в задней, цилиарной зоне |
радужной оболочки. G помощью |
|||||
мышц регулируется |
поступление |
лучей света в глазное яблоко, |
||||
то есть |
радужная |
оболочка |
выполняет |
функцию диафрагмы. |
||
Мышца, |
расширяющая |
зрачок, |
иннервируется постганглиопар- |
|||
ными симпатическими |
волокнами краниального шейного узла, а |
|||||
сфинктер |
зрачка — постганглионариыми парасимпатическими во |
|||||
локнами |
цилиарного узла. |
|
|
|
||
У лошади и жвачных у свободного зрачкового края радужной оболочки имеются в ы р о с т ы (з е р н а), пронизанные кровенос ными сосудами и содержащие сильно пигментированные клетки.
Ресничное (цилкарное) тело (corpus ciliare) — утолщенная
часть средней оболочки, расположенная между радужной и сосу
дистой оболочками. Различают заднюю, более |
тонкую |
часть с |
|
мелкими складками — р е с н и ч н о е |
к о л ь ц о |
и переднюю, бо |
|
лее толстую с высокими отростками, |
направленными к |
хруста |
|
лику,— р е с н и ч н ы й в е н ч и к (цилиариая корона). Отростки и складки цилиарного тела покрыты цилиарной частью сетчатки — эпителием, имеющим два слоя: наружный из пигментированных клеток и внутренний из клеток, лишенных пигмента, обращен ных к полости глаза. Эпителиальные клетки принимают участие- в образовании жидкости, заполняющей переднюю и заднюю ка меры глаза. Основная масса цилиарного тела состоит из реснич ной мышцы, образованной пучками гладких мышечных клеток, расположенных в трех направлениях: кольцевом, радиальном и меридиональном. Между мышечными пучками расположена со единительная ткань, содержащая кровеносные капилляры и пиг ментные клетки.
Благодаря двигательной активности мышц ресничное тела имеет большое значение в аккомодации глаза. При сокращении мышц натяжение связки, поддерживающей хрусталик, ослабля ется, и он становится более округлым, что приспосабливает глаз к рассматриванию предметов, находящихся на близком расстоя нии. При расслаблении мышц достигается противоположный эффект.
Сосудистая оболочка |
(tunica vasculosa) — задняя часть сред |
ней оболочки, отличается |
обилием кровеносных сосудов. Состоит |
из соединительной ткани, в которой развита сеть эластических волокон и много пигментных клеток. В соответствии со строе нием в оболочке различают четыре пластипки: надсосудистую, сосудистую, хориокапиллярную и базальную. С помощью надсосудистой пластинки сосудистая оболочка соединяется со склерой, в сосудистой пластинке содержится сеть крупных сосудов, а в \ ориокапиллярной — густая сеть кровеносных капилляров. Между сосудистой и хориокапиллярной пластинками у животных распо ложена бессосудистая зона, состоящая из многогранных клеток у плотоядных (tapetum lucidum) или из переплетающихся соеди нительнотканных волокон (tapetum fibrosum). У кошки в цито плазме клеток тапетума имеются правильно расположенные игольчатые кристаллы. Считают, что наличием этого слоя обус ловлено свечение глаз в темноте отраясенным светом. Базальиои пластинкой сосудистая оболочка отделена от пигментного эпите лия сетчатки.
С е т ч а т к а |
(retina) — внутренняя |
оболочка |
стенки |
глазного |
|||||
яблока, прилежащая к стекловидному телу. |
В соответствии с рас |
||||||||
положением, строением и |
функцией в сетчатке |
различают две |
|||||||
части: |
н е р в н о к л е т о ч н у ю |
(pars |
nervosa) |
з р и т е л ь н у ю , |
|||||
выстилающую |
изнутри заднюю, |
большую |
часть |
стенки |
глазного |
||||
яблока, |
и п е р е д н ю ю |
п и г м е н т н у ю |
(pars |
pigmentosa), по |
|||||
крывающую изнутри |
ресничное |
тело |
и |
радужную |
оболочку. |
||||
В зрительной |
части |
происходит |
восприятие световых |
раздраже- |
|||||
^49 |
16* |
24Э |
|
|