

gistologia
.pdfП р е д п л о д н ы й п е р и о д начинается с конца 8-го дня и завершается к 13 суткам инкубации. Все провизорные органы до стигают функциональной зрелости. Наряду с основным внекишечным способом питания зародыша, который осуществляется посред ством желточного мешка, имеет место усвоение материала внутрикишечным способом: цыпленок заглатывает амниотическую жидкость из полости амниона.
Дыхание предплода осуществляется исключительно через сосудистую систему аллантоиса.
П л о д н ы й п е р и о д охватывает срок с 13—14 суток до на чала 20-х суток инкубации. Характерным для этого периода явля ется интенсивный рост органов тела плода. Основной способ питания зародыша — это внутрикишечный, внекишечный способ становится дополнительным. Белок, переместившийся в полость амниона и смешавшийся с амниотической жидкостью, является источником питания. Сохраняется аллантоидное дыхание. Крове носные сосуды аллантоиса находятся вблизи подскорлуповой обо лочки. Это способствует усилению газообмена, что необходимо в связи с интенсивным развитием органов тела цыпленка.
П е р и о д в ы л у п л е н и я — на 20—21-е сутки инкубации. "Он характеризуется прекращением функций временных органов, кишечным пищеварением, дыханием с участием легких.
Таким образом, данные о приспособительных реакциях орга низма и о закономерностях формообразования, свойственные каж дому периоду эмбриогенеза, являются научным обоснованием при разработке и внедрении мероприятий, направленных на дальней шее совершенствование технологии искусственного выведения •сельскохозяйственных птиц.
ЭМБРИОНАЛЬНОЕ РАЗВИТИЕ МЛЕКОПИТАЮЩИХ
В характеристике развития млекопитающих будут освещены во просы, касающиеся строения половых клеток, оплодотворения, особенности дробления, образования гаструлы, дифференцировки зародышевых листков и осевых органов, развития, строения и -функции плодных оболочек (провизорных, или временных, орга нов).
Подтип млекопитающих по характеру эмбриогенеза очень раз нообразен. Усложнение строения млекопитающих, а следователь но, и эмбриогенеза обусловливает необходимость накопления большего количества питательных вешеств в яйцеклетках. На определенном этапе развития этот запас питательных веществ не может удовлетворить потребности качественно изменепного заро дыша, в связи с чем в процессе эволюции у млекопитающих выра боталось внутриутробное развитие и у, большинства животных этого подтипа наблюдается вторичная потеря яйцеклетками желтка.
Половые клетки. Оплодотворение. Дробление. Самыми прими тивными млекопитающими являются яйцекладущие (утконос,
ехидна). У них телолецитальные яйца, меробластическое дробле* иие, поэтому их эмбриогенез сходен с развитием птиц.
У сумчатых млекопитающих я й ц е к л е т к и содержат незна чительное количество желтка, но зародыш рождается недоразви тым и дальнейшее его развитие протекает в материнской сумке, где устанавливается связь соска молочной железы матери с пище водом детеныша.
Для высших млекопитающих характерно внутриутробное раз витие и питание зародыша за счет материнского организма, что и отразилось на эмбриогенезе. Яйцеклетки почти полностью вто рично утратили желток; их считают вторично олиголецитальными, изолецитальными. Они развиваются в фолликулах (folliculus — мешочек, пузырек) яичника. После овуляции (разрыва стенки фолликула и выхода яйцеклетки из яичника) они попадают в яйцевод. •
Яйцеклетки у млекопитающих микроскопических размеров. Их диаметр равен 100—200 мкм. Они покрыты двумя оболочками — первичной и вторичной. Первая — это плазмолемма клетки. Вто рой оболочкой являются фолликулярные клетки (см. рис. 37). Из= них построена стенка фолликула, где находятся яйцеклетки в яичнике.
О п л о д о т в о р е н и е яйцеклетки протекает в верхней части яйцевода. При этом оболочки яйцеклетки разрушаются под воз действием ферментов акросомы спермия.
Д р о б л е н и е у высших млекопитающих полное, асинхрон ное: образуется зародыш, состоящий из 3, 5, 7 и т. д. бластомеров.. Последние обычно лежат в виде кучки клеток. Эта стадия носит название морулы (рис. 62). В ней различимы два типа клеток: мелкие — светлые и крупные — темные. Наибольшей митотической активностью обладают светлые клетки. Интенсивно делясь, они располагаются на поверхности морулы в виде наружного слоя трофобласта (trophe — питание, blastos — росток). Темные бластомеры делятся медленней, поэтому они крупнее светлых и нахо дятся внутри зародыша. Из темных клеток образуется эмбриобласт.
Трофобласт выполняет трофическую функцию. Он обеспечи вает зародыш питательным материалом, так как с его участием устанавливается связь зародыша со стенкой матки. Эмбриобласт— это источник развития тела зародыша и его некоторых внезародышевых органов.
Если у животных рождается несколько детенышей, то в яйцепод поступает сразу несколько яйцеклеток.
Дробясь, зародыш продвигается по яйцеводу в направлении матки (рис. 63, 64). Трофобласт впитывает секрет желез. Он скап ливается между эмбриобластом и трофобластом. Зародыш сильно увеличивается в размере и превращается в бластодермический пузырек, или бластоцисту (рис. 65). Стенкой бластоцисты являет ся трофобласт, а эмбриобласт имеет вид кучки клеток и называ ется зародышевым узелком.
82
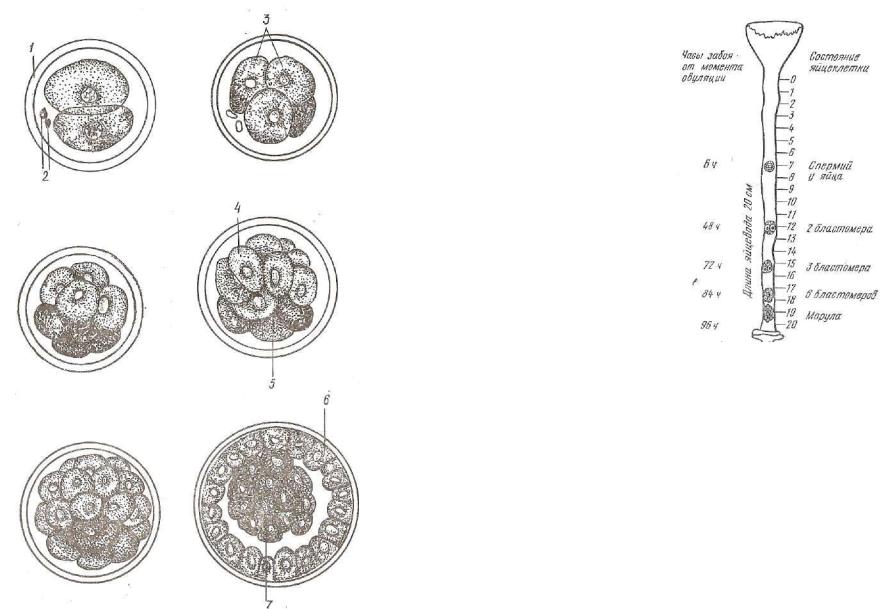
Рис. 62. Схема дробления яйца млекопитающего:
1 — блестящая оболочка; 2 — полярные тельца; 3 — бластомеры; 4 — светлые бластомеры, образующие трофобласт; 5 — темные бла стомеры; 6 — трофобласт; 7 — зародышевый узелок.
Рис. 63. Схема перемещения дробя щейся зиготы коровы по яйцеводу.
Полость бластоцисты |
запол |
|
||
нена жидкостью. Она образова |
|
|||
лась |
вследствие |
поглощения |
|
|
клетками трофобласта |
секрета |
|
||
маточных желез. |
Вначале бла- |
|
||
стоциста находится свободно в |
6v |
|||
полости матки. Затем при помо |
|
|||
щи ворсинок, образовавшихся на |
|
|||
поверхности трофобласта, бла- |
|
|||
стоциста прикрепляется к стен- |
^<?у |
|||
ке матки. Этот процесс называ |
|
|||
ется имплантацией (im — про |
|
|||
никновение в, plantatio — посад- |
72 ч |
|||
ка) (рис. 66). У крупного рога- ? |
|
|||
того скота имплантация насту- |
8$ ч |
|||
пает на 17-е сутки, у лошади на |
|
|||
63—70-е сутки, у макаки — на |
двч |
|||
9-й день после оплодотворения. |
|
|||
Затем |
клетки |
зародышевого |
|
|
узелка выстраиваются в виде пласта — формируется зародышевый диск, аналогичный зародышевому диску птиц. В его средней части дифференцируется уплотненная зона — зародышевый щпток. Как и у птиц, из материала зародышевого щитка развивается тело за родыша, а остальная часть зародышевого диска используется при образовании провизорных органов.
Таким образом, несмотря на то что у высших млекопитающих из-за вторичной потери желтка яйцеклетки относятся к олиголецитальным с голобластическим дроблением, строение бластулы сходно с такой, которая образуется после меробластического дробления. Это можно объяснить тем, что предшественники мле копитающих имели полилецитальные, телолецитальпые яйца и высшие млекопитающие унаследовали от своих предков строение бластулы, последняя напоминает бластулу птиц.
Гаструляция. Закладка осевых органов и их дифференциация. Гаструляция протекает так же, как у пресмыкающихся, птиц, низ ших млекопитающих. Путем деляминации зародышевого диска формируются эктодерма и энтодерма. Если эти листки образова лись из материала зародышевого щитка, то они называются заро дышевыми, а если они возникли из незародышевой зоны зароды шевого диска, то незародышевыми. Незародышевые эктодерма и энтодерма разрастаются по внутренней поверхности трофобласта. Вскоре трофобласт, расположенный над зародышем, рассасываотся и последний оказывается некоторое время лежащим в поло сти матки ничем не прикрытый.
85
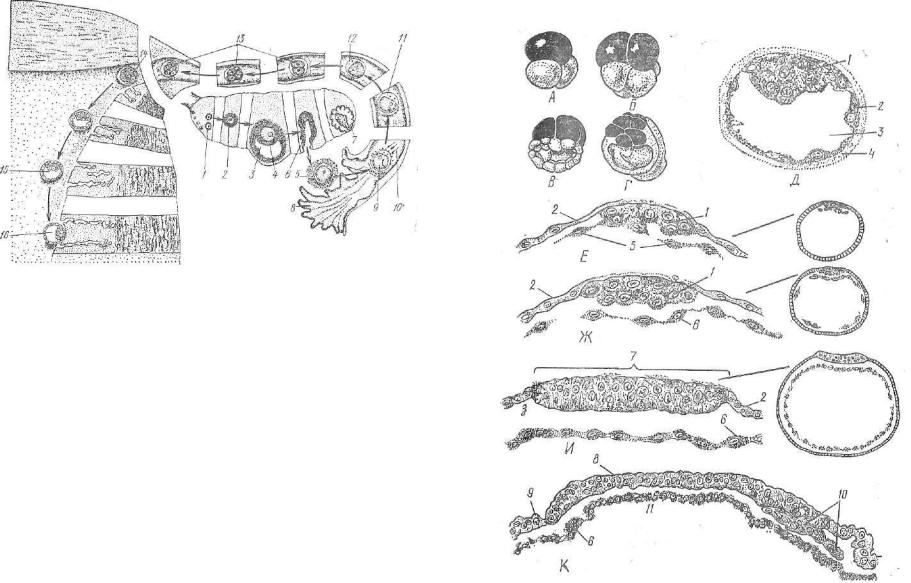
Рис. 64. Схема овуляции, оплодотворения, дробления, имплантации:
1 — примордиальные фолликулы; 2 — растущие фолликулы; 3, 4 — пузырчатые* фолликулы; 5 — овулировавшая яйцеклетка; 6 — спавшийся пузырчатый фолли кул; 7 — желтое тело; * — фимбрии воронки яйцевода; 9 — яйцеклетка в моментпроникновения в нее спермин; Ю — спермин; 11 — зигота, сближение пронуклеусов; 12 — зигота в метафазе; 13 — дробление; 14 — морула; 15 — бластоциста; 16 — имплантация.
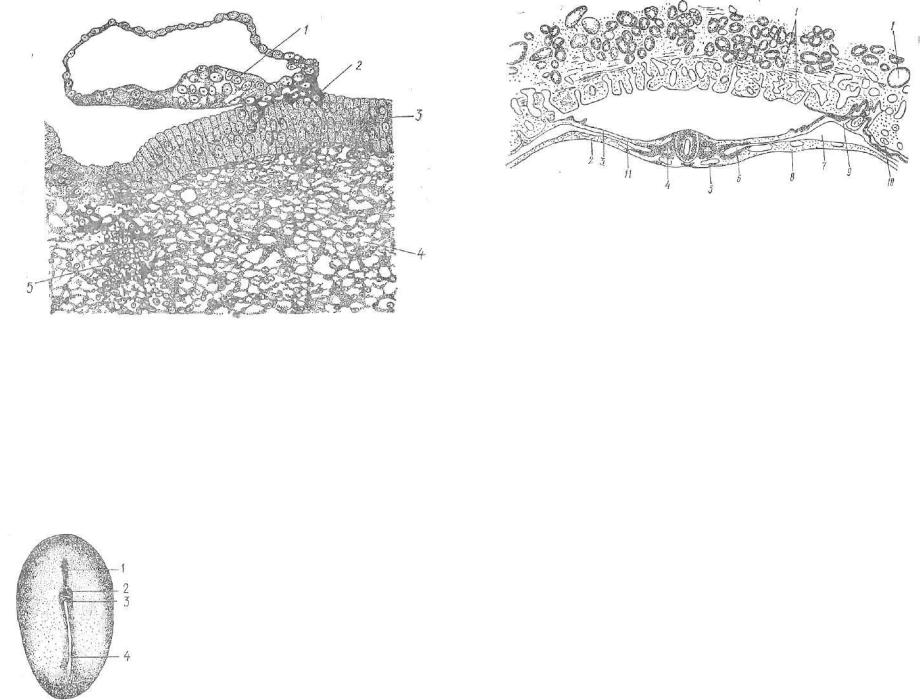
Формирование мезодермы протекает так же, как и у птиц; Клетки краевой зоны дискобластулы мигрируют двумя потоками к задней части зародыша. Здесь эти потоки встречаются и изме няют свое направление движения. Теперь они перемещаются впе ред в центре зародышевого диска, формируя при этом первичнуюполоску с продольным углублением — первичной бороздкой. На переднем конце первичной полоски образуется гензеновский узе лок с углублением — первичной ямкой. В этой зоне подворачива ется материал будущей хорды и растет вперед между эктодермой и энтодермой в виде головного (хордалыюго) отростка (рис. 67).
Из клеток первичной полоски развивается мезодерма. После миграции ее материал растет между эктодермой и энтодермой и превращается в сегментированную мезодерму (сомиты), прилега ющие к пей сегментные ножки и несегментированную мезодерму. Сомиты состоят из склеротома (вентромедиальная часть), дермотома (латеральная часть), миотома (медиальная часть). Сомитымогут соединяться с несегментированной мезодермой посредством сегментных пожек. Несегментировапиая часть мезодермы имеет вид полого мешка. Его наружная стенка называется париеталь ным листком, ft внутренняя — висцеральным. Полость, заключен ная между ними, именуется вторичной полостью тела, или целомом (рис. 68).
Л—Г — последовательные стадии дробления (черным — бластомеры, из которых ппзовьется тело зародыша; 1 белым — бластомеры, из которых разовьется трофоОласт); Д — бластоциста; Е—И — развитие зародышевого диска и образование эн тодермы; К — образование мезодермы и первичной кишки из энтодермы; 1 — за родышевый узелок; 2 — трофобласт; з — бластоцель; 4 — блестящая зона; 5 — клетки энтодермы; б — энтодерма; 7 — зародышевый диск; н — эктодерма sapoдишевого диска; 9 — трофэктодерма; ю — мезодерма; 11 — первичная кишка (стенка) (по Пэттену).
86
Дпфференцировка зародышевых листков протекает так жег как у птиц и других животных. На спинной части зародыша в эктодерме образуется нервная пластинка; после срастания ее кра ев формируется нервная трубка. На нее нарастает эктодерма, по этому очень скоро нервная трубка оказывается погруженной под эктодерму. Из нервной трубки развивается вся нервная система, из эктодермы — поверхностный слой кожи (эпидермис). Хорда как орган у взрослых животных не функционирует. Она полно
стью замещается позвонками позвоночного столба. Миотомы сомитов являются источни ком формирования мышц туловища, а склеротомы — мезенхимы, из которой затем разви ваются костная и хрящевая ткани. Дерматом — зачаток глубоких слоев кожного покро-
Рис. 67. Зародыш кролика, вид сверху:
1 — головной отросток; 2 — гензеновский узелок; 3 — первичная ямка; 4 — первичная полоска.
2 — железы матки; 2 — висцеральный из — париетальный листки мезодермы; 4— миотом; 5 — аорта; в — внутризародышевый целом; 7 — внезародышевый целом; 8 — энтодерма желточного мешка; 9 — ворсинки хориона; 10 — трофобласт; 11— энтодерма.
ва. Из материала сегментных ножек образуются мочевыделительиая и половая системы, поэтому его называют нефрогонадотомом.
Из париетального листка спланхнотома формируется поверх ностная ткань (эпителий) париетального листка плевры и брю шины, из висцерального листка — эпителий серозных оболочек тех органов, которые лежат в грудной и брюшной полостях.
Из энтодермы развивается эпителий, покрывающий внутренюю поверхность пищеварительной трубки и органы — производные пищеварительной трубки: органы дыхания, печень, поджелудоч ная железа.
Таким образом, развитие зародышевых листков и их дальней шая дифференциация у млекопитающих сходны с таковыми у дру гих животных. Эти признаки являются наиболее древними, в них отражен путь, который прошли в своём развитии млекопитающие. Такие признаки относят к палингенетическим (palin — снова, ge nesis— рождение) в отличие от ценогенетических, то есть приоб ретенных в связи с изменениями условий обитания, например выходом животных из воды на сушу.
Из зародышевых листков — эктодермы, энтодермы и мезодер мы развиваются не только постоянные органы зародыша. Они участвуют в закладке временных, или провизорных, органов — плодных оболочек.
Образование внезародышевых (временных) органов (рис. 69). Одной из особенностей развития млекопитающих считают, что при и80лецитальной яйцеклетке и голобластическом дроблении про исходит образование временных органов. Как известно, в эволю ции хордовых провизорные органы — это приобретение позвоноч ных с телолецитальными, полилецитальными яйцами и меробласиическим дроблением.
89
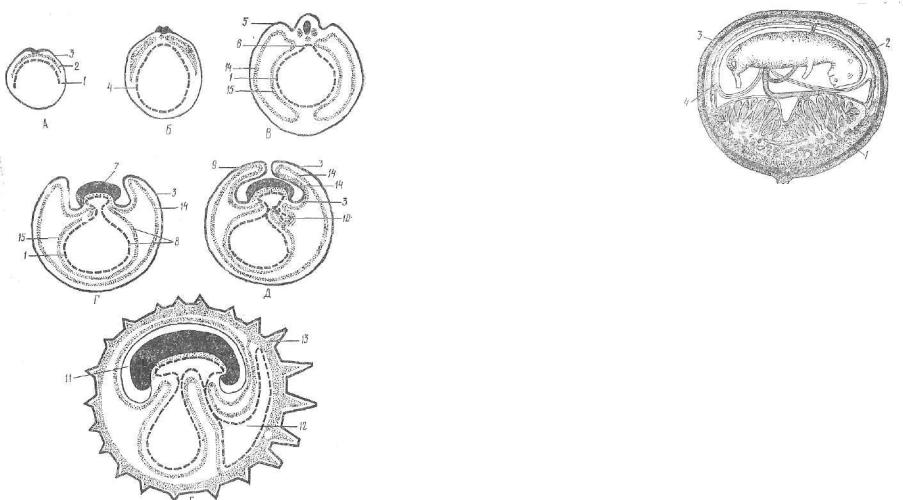
Рис. 69. Схема развития желточного мешка и зародышевых оболочек у мле копитающих (шесть последовательных стадий):
А — процесс обрастания полости плодного пузыря энтодермой (1) и мезодермой (2); Б — образование замкнутого энтодермального пузырька (4); В — начало об разования амниотической складки (5) и кишечного желобка (в); Г — обособление тела зародыша И); желточный мешок (8); Д — смыкание амниотических складок (9); начало образования развития аллаитоиса (10); Е — замкнутая амниотическаяполость (11); развитый аллантоис (12); ворсинки хориона (13); париетальный ли сток мезодермы (14); висцеральный листок мезодермы (15); эктодерма (3).
Другая особенность развития млекопитающих — это очень раннее обособление зародышевой от незародышевой части. Так,. уже в начале дробления образуются бластомеры, формирующие виезародышевую вспомогательную оболочку — трофобласт, с по мощью которого зародыш начинает получать питательные веще-
90
Рис. 70. Схема взаимоотношений мат ки и желточного мешка у кролика:
1 — аллантслвдная плацента; 2 — жел точный мешок; 3 — стенка матки; 4 — амнион.
ства из полости матки. После образования зародышевых лист ков трофобласт, расположенный над зародышем, редуцируется. Нередуцированная часть трофобласта, срастаясь с эктодермой, образует единый слой. Прилегая
с внутренней стороны к этому слою, растут листки несегментироианяой мезодермы и впезародышевая эктодерма.
Одновременно с формированием тела зародыша протекает раз витие плодных оболочек: желточного мешка, амниона, хориопа, зллантоиса.
Ж е л т о ч н ы й м е ш о к , как и у птиц, образуется из внезародышевых энтодермы и висцерального листка мезодермы. В от
личие от |
птиц он содержит не желток, а |
белковую жидкость. |
В стенке |
желточного мешка формируются |
кровеносные сосуды. |
Эта оболочка выполняет функции кроветворения и трофическую •функцию. Последняя сводится к переработке и доставке питатель ных веществ от материнского организма к зародышу (рис. 70,71). Продолжительность функции желточного мешка у разных жи вотных различна.
Как и у птиц, у млекопитающих развитие плодных оболочек начинается с образования двух складок — туловищной и амнио тической. Туловищная складка приподнимает зародыш над жел точным мешком и отделяет его зародышевую часть от незароды шевой, а зародышевая энтодерма смыкается в кишечную трубку. Однако кишечная трубка остается связанной с желточным меш ком узким желточным стебельком (протоком). Острие туловищ ной складки направлено под туловище зародыша, при этом проги баются все зародышевые листки: эктодерма, несегментированпая мезодерма, энтодерма.
В образовании амниотической складки участвует трофобласт, сросшийся с внезародышевой эктодермой и париетальным лист ком мезедермы. В амниотической складке имеются две части: внутренняя и наружная. Каждая из них построена из одноимен ных листков, но отличается порядком их расположения. Так, внутренним слоем внутренней части амниотической складки явля ется эктодерма, которая в наружной части амниотической склад ки будет находиться снаружи. Это касается и последовательно сти залегания париетального листка мезодермы. Амниотическая складка направлена над телом зародыша. После срастания ее краев эародыш становится окруженным сразу двумя плодными
оболочками — амнионом и хорионом. |
_, |
|
91 |

Рис. 71. Схема миграции первичных половых клеток из желточного мешка в зачаток гонады (разяв1е этапы миграции условно нанесены на один и тог же поперечный срез зародыша):
1 — эпителий желточного мешка; 2 — мезенхима; 3 — сосуды; 4 — первичная почка; 5 — зачаток гонады; 6 — первичные половые клетки; 7 — зачатковый эпи телий.
А м н и о н развивается из внутренней части . амииотической складки, хорион — из наружной части. Полость, которая образо валась вокруг зародыша, называется амииотической полостью. Она заполнена прозрачной водянистой жидкостью, в образовании которой принимают участие амнион и зародыш. Амниотическая жидкость предохраняет зародыш от излишней потери воды, слу жит защитной средой, смягчает удары, создает возможность подвижности зародыша, обеспечивает обмен околоплодных вод. Стенка амниона состоит из внезародышевой эктодермы, направ ленной в полость амниона и расположенного снаружи эктодермы париетального листка мезодермы.
Х о р и о н гомологичен серозе птиц и других животных. Он развивается из наружной части амииотической складки, а поэто му построен из трофобласта, соединенного с эктодермой, и парие тального листка мезодермы. На поверхности хориона образуются отростки — вторичные ворсинки, врастающие в стенку матки. Эта зона сильно утолщена, обильно снабжена кровеносными сосуда ми и называется детским местом, или плацентой. Основной функ цией плаценты является снабжение зародыша питательными веществами, кислородом и освобождение его крови от углекисло ты и ненужных продуктов обмена. Поступление веществ в кровь зародыша и из нее осуществляется диффузным путем или с по мощью активного переноса, то есть с затратой на этот процесс
Рис. 72. Схема взаимоотношений ор ганов у плода животных с эпителиохориальным типом плацентации:
1 — алланто-амнион; 2 — алланто-хори- 2 он; 3 — ворсинки хориона; 4 — полость мочевого мешка; 5 — полость амниона; / - 6 — желточный мешок.
энергии. Следует, однако, обра тить внимание на то, что кровь матери ни в зоне плаценты, ни в других участках хориона не смешивается с кровью плода.
П л а ц е н т а , являясь органом питания, выделения, дыхания плода, выполняет и функцию органа эндокринной системы. Гор моны, синтезируемые трофобластом, а затем плацентой, обеспечи вают нормальное течение беременности.
По форме различают несколько типов плаценты.
1. Диффузная плацента (рис. 72) — вторичные сосочки ее развиваются по всей поверхности хориона. Встречается она у свиньи, лошади, верблюда, сумчатых, китообразных, бегемота. Ворсинки хориона проникают в железы стенки матки, не разру шая при этом ткани матки. Так как последняя покрыта эпители ем, то по строению такой тип плаценты называют эпителиохориальной, или полуплацентой (рис. 73). Питание зародыша осуще ствляется следующим способом — маточные железы секретируют маточное молочко, оно всасывается в кровеносные сосуды ворси нок хориона. При родах ворсинки хориона выдвигаются из маточ ных желез без разрушения тканей, поэтому кровотечение при этом обычно отсутствует.
2. Котиледонная плацента (рис. 74) —ворсинки хориона рас-- положены кустиками — котелидонами. Они соединяются с утол щениями стенки матки, которые именуются к а р у н к у л а м и . Комплекс котиледон—карункул называется плацентомом. В этой1 зоне эпителий стенки матки растворяется и котиледоны погру жены в более глубокий (соединительнотканный) слой стенки мат ки. Такая плацента называется десмохориальной и свойственна парнокопытным. По мнению некоторых ученых, и у жвачных — плацента эпителиохориальная.
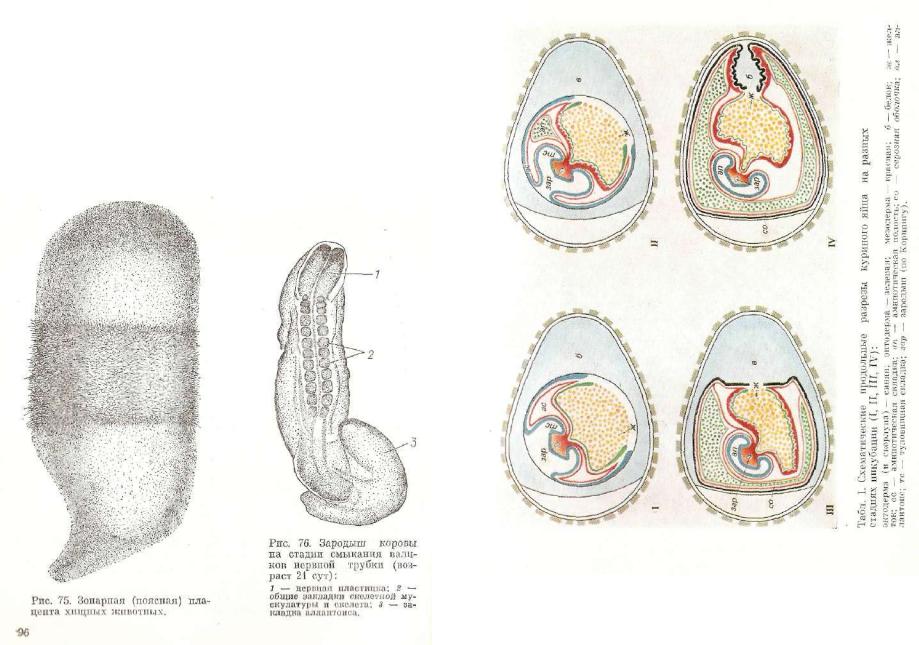
3.Поясная плацента (рис. 75). Зона залегания ворсинок хо риона в виде широкого пояса окружает плодный пузырь. Связь зародыша со стенкой матки более тесная: ворсинки хориона рас полагаются в соединительнотканном слое стенки матки, контакти руя с эндотелиальным слоем стенки кровеносных сосудов. Эта плацента называется эндотелиохориальной.
4.Дискоидальная плацента. Зона контакта ворсинок хориона
истенки матки имеет форму диска. Ворсинки хориона погружа ются в заполненные кровью лакуны, лежащие в соединительно-
Ь2 |
93. |

тканном слое стенки матки. Такой тип плаценты называется гемохориальпой и встречается у приматов.
А л л а н т о и с — вырост вентральной стенки задней кишки. Как и кишка, он состоит из энтодермы и висцерального листка мезодермы. У некоторых млекопитающих в нем скапливаются азотистые продукты метаболизма, поэтому он функционирует как мочевой пузырь. У большинства животных в связи с очень ранним развитием зародыша с материнским организмом аллантоис развит значительно слабее, чем у птиц. Через стенку аллантоиса прохо дят кровеносные сосуды от эмбриона и плаценты. После врастания кровеносных сосудов в аллантоис последний начинает принимать участие в обмене веществ зародыша.
Место соединения аллантоиса с хорионом называется хориоаллантоисом или аллантоидной плацентой. Связь зародыша с пла центой осуществляется посредством пупочного канатика. В его •состав входят узкий проток желточного мешка, аллантоис п кро-
а — эпителиохориальная; б— десмохориальная; в — эндотелиохориальная; г —« гемохориальная; 1 — эпителий хориона; 2 — эпителий стенки матки; 3 — соедич аительная ткань ворсинки хориона; 4 — соединительная ткань стенки матки; 5— кровеносные сосуды ворсинок хориона; 6 — кровеносные сосуды стенки матки; 7— материнская кровь.
веносные сосуды. У некоторых животных с плацентой связан и желточный мешок. Такая плацента называется желточной.
Таким образом, продолжительность эмбриогенеза у разных пла центарных животных различна. Она обусловлена зрелостью рожде ния детенышей и характером связи зародыша с организмом мате ри, то есть строением плаценты.
Эмбриогенез сельскохозяйственных животных протекает ана логично и отличается от приматов. Эти особенности развития бу дут кратко освещены ниже.
В акушерской практике внутриутробное развитие делят на три периода: эмбриональный (зародышевый), предплодный и плод ный. Зародышевый период характеризуется развитием признаков,, типичных для всех позвоночных и млекопитающих. В предплод ный период закладываются признаки, свойственные данному се мейству. В плодпый период развиваются видовые, породные и ин дивидуальные особенности строения.
У крупного рогатого скота продолжительность внутриутроб ного развития 270 дней (9 мес). По данным Г. А. Шмидта, заро-
95
дышевый (эмбриональный) период длится первые 34 дня, преднлодный период — с 35-го по 60-й день, плодный период — с 61-го по 270-й день.
В течение первой недели протекает дробление зиготы и обра зование трофобласта. Питание зародыша осуществляется за счет желтка яйцеклетки. При этом идет безкислородное расщепление питательных веществ.
С 8-х по 20-е сутки — это стадия развития зародышевых лист ков, осевых органов, амниона и желточного мешка (рис. 76). Пи тание и дыхание осуществляются, как правило, с помощью трофо- •б ласт а.
На 20—23-е сутки развивается туловищная складка, формиру ется пищеварительная трубка и аллантоис. Питание и дыхание протекают с участием кровеносных сосудов.
24—34-е сутки — стадия образования плаценты, котиледонов хориона, многих систем органов. Питание и дыхание зародыша
кп; |
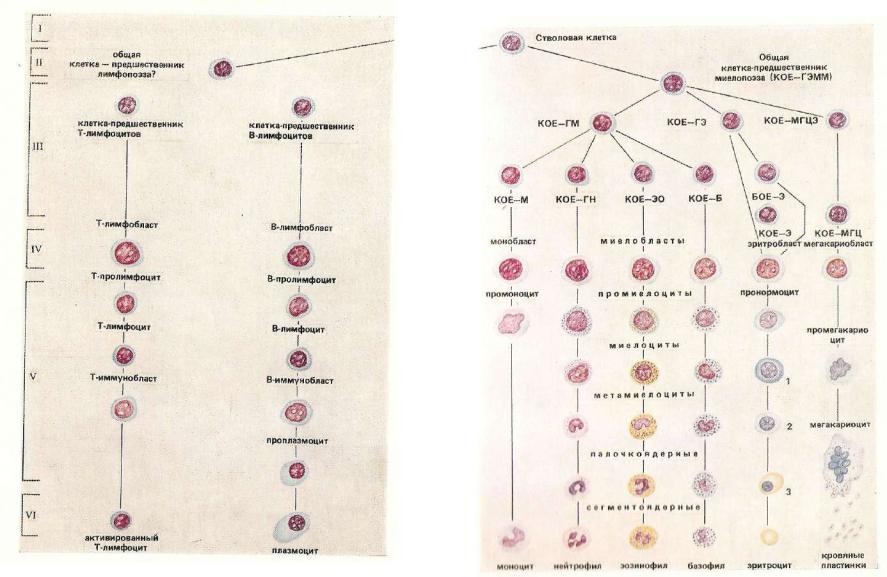
II — полустволовые, частично детермини- |
ну, 1081): |
|
— |
бласты; V — созревающие клетки: VI - |
||
роваииыв полшттоцтныо IUI отки-н р о дш сотне ни ик и миелопоэза и лимфопоэза; III— |
|||
|
|
||
|
|
зрелые клетки; 1 — ЛавофилышН нормоцит; 2 — полихроматофильный нормоцит; |
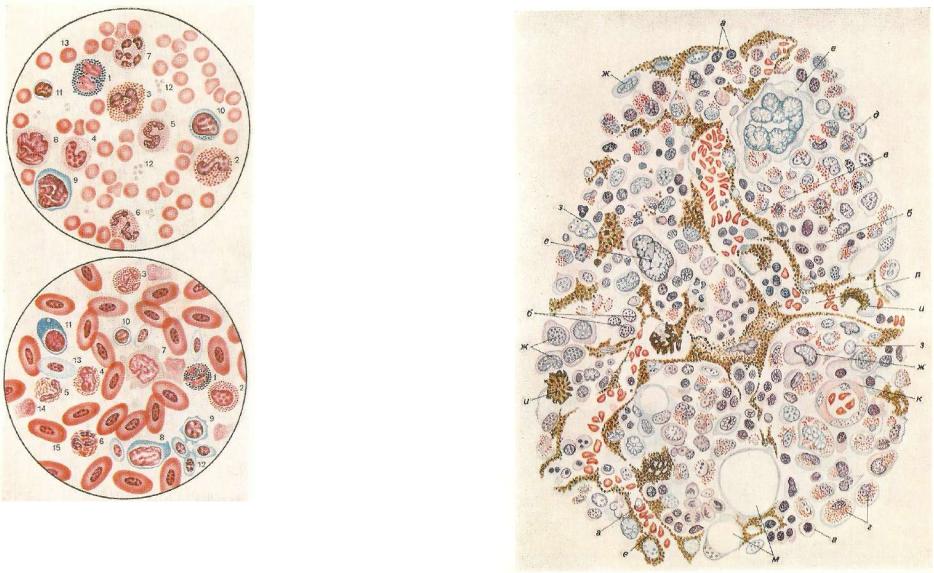
Табл. IV. Форменные эле менты крови курицы в ок рашенном мазке (по Ники тину):
1—• палочкоядерный базофил;
2— эозинофильный миелоцит;
3— сегментоядерный эозино фил; 4, 5, в — нейтрофилы, псевдоэозинофилы; (4 — мие лоцит; 5 — палочкоядерный;
б— сегментоядерный); 7 — моноцит; 8 — большой, о — средний, ю — малый лимфо циты; а — плазмоцит; 12 — тромбоциты; 13 — полихроматофильный эритроцит; 14 — остатки поврежденного эрит роцита; is — овальные ядер ные эритроциты.
Табл. V. Схема строения красного костного мозга (по Максимову):
и — нормоциты; б — эритробласты; в —.эритроциты; г — аозииофильные миелоциты; в — эозинофильные метамиелоциты; ед§г мегакариоциты; ж — миелобласты; з — ретикулярные клетки; и — макрофаги; к — артериальный капилляр; л — ве нозный синусоид; м — жировые клетки.