

gistologia
.pdfции — пассивный перенос, так и с затратами энергии против гра диента концентраций — активный перенос.
В состав оболочки входят плазматическая мембрана, надмембранный комплекс •— гликокалекс и субмембранный опорпо-сокра- тительный аппарат. В основе плазматической мембраны лежит •бимолекулярный слой липидов, в который погружены полностью яли частично молекулы белка и гликопротеидов. В ней около 40% липидов, 60% белков и до 1% углеводов. В связи с функциональ ной характеристикой клеток различных тканей специфичен состав гликопротеидного надмембранного комплекса. В нем содержится до 1% углеводов (гиалуроновая, сиаловая кислоты и др.), молеку лы которых образуют длинные ветвящиеся цепи полисахаридов, связанные с белками мембраны (рис. 12 и 13). Находящиеся в гликокалексе белки — ферменты участвуют в конечном внеклеточ ном расщеплении веществ. Продукты этих реакций в виде мономе ров поступают в клетку. При' активном переносе транспорт ве ществ в клетку (эндоцитоз) осуществляется или поступлением молекул в виде раствора — пипоцитоз, или захватом крупных частиц — фагоцитоз.
Процесс ф а г о ц и т о з а состоит из двух фаз: взаимодействие частицы с рецептором плазмолеммы клетки и затем поглощение ее в результате образования псевдоподий. Первоначальное взаимо действие частицы и рецептора плазмолеммы вызывает сигнал, в результате которого происходят местные скопления контрактильных белков (актина и др.) в поверхностном слое цитоплазмы, ве дущие к образованию псевдоподий. Это увеличивает площадь ее контакта с частицей, что вызывает дальнейшее скопление сокра тимых белков. Процесс продолжается, пока псевдоподии не сом кнутся над частицей, формируя фагосому.
П и н о ц и т о з — везикулярное поглощение жидкости, содер жащей низкомолекулярные растворы (липопротеиды, иммунные комплексы, ферритин, гормоны и др.)- Различают макропиноцитоз, при котором ундулирующие складки поверхности клетки за хватывают капли раствора, видимые при фазоконтрастной микро скопии, и микропиноцитоз — жидкость захватывается минималь ными инвагинациями, различимыми только при электронной микроскопии.
По механизму действия микропиноцитоз бывает жидкофазный и абсорбтивный. Первый — неизбирательный: растворенные веще ства поглощаются пропорционально их концентрации в жидкой среде, а поглощающая их мембрана морфологически не специали зирована. При втором — мембраны пузырькообразпых инвагина ций плазмолеммы клетки покрыты с внешней поверхности тонким местным слоем гликокалекса, а с внутренней — щетинкой тонких волосков.
Количество интериоризированной (погруженной) при эндоцитозе мембраны может быть большим, особенно при фагоцитозе. Макрофаги in vitro могут интериоризировать при фагоцитозе до 18% своей плазмолеммы в час.
|
|
Цитоплазма |
|
^ Л п п ' |
я |
С Д е ^ ^ Р О е И Е Я п л а з м а ™ ч е с к о ? мембраны: |
белки с внешней сторо |
ны слоя связаны с полисахаридами, образуя слой |
гликокалекса. |
||
20
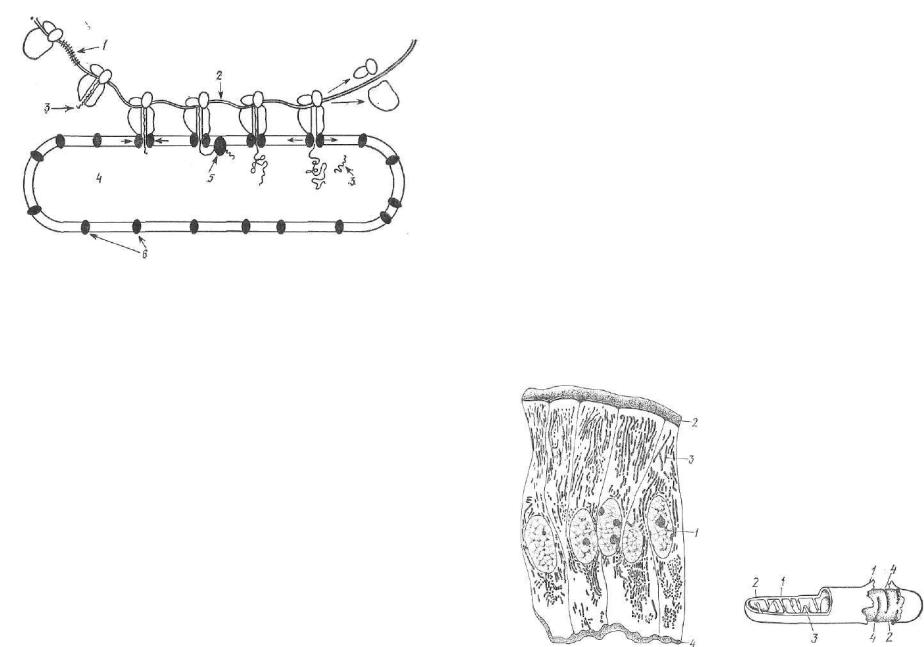
Рис. 14. Схема клеточных контактов:
I — простой контакт; 2 — вамок; з — плотный контакт; 4 — промежуточный контакт; 5 — десмосома; б — ще левой контакт.
В соответствии с функциональными и морфо логическими особенностями тканей оболочка клеток образует характерные для них аппа раты м е ж к л е т о ч н ы х к о н т а к т о в . Ос новные их формы: простой контакт, плотный контакт, промежуточный контакт (или' зона слипания) и щелевой контакт (рис. 14).
Простой контакт — наиболее распростра ненная форма контакта двух смежных клеток. При нем клетки отстоят одна от другой на расстоянии 15—20 нм. Межклеточное прост ранство соответствует надмембранным компо нентам клеточных мембран контактирующих клеток.
Плотный (замыкающий) контакт. При нем впепшие слои плазмолеммы у люминальной поверхности смежных клеток сливаются в одну общую структуру и изолируют межкле точное пространство от внешней для ткани среды. Этот тип соединения находится между эпителиальными клетками у их апикальной
поверхности и образует зону слияния мембран (слипания их ин тегральных белков), окружающую в виде пояска верхушки клеток. Белки мембраны связаны в зоне плотного замыкающего контакта с системой тонких фибрилл цитоплазмы, ориентированных парал лельно поверхности клетки по ходу зоны слипания.
Разновидностью плотного контакта являются десмосомы. Они характеризуются особым развитием и дифференцировкой надмембранного комплекса смежных клеток. В точечных десмосомах расстояние между мембранами двух контактирующих клеток 22—35 нм. В межклеточном пространстве за счет надмембранного комплекса формируется волокнистое вещество. В его центральной части образуется пластинка, содержащая белки и мукополисахариды. Она связана с плазмолеммами смежных клеток поперечными фибриллами. К мембранам контактирующих клеток прилегают электроноплотные зоны цитоплазмы с отходящими от них фибрил лами. Десмосомы обеспечивают механическую связь смежных клеток.
Щелевой контакт характеризуется наличием незначительного межклеточного пространства (до 2—3 нм). Это специализирован ная область плазмолемм смежных клеток, обеспечивающая диффу зию ионов и мелких молекул от одной клетки в другую. При со ответствующей обработке ткани электроноплотным веществом видно, что межклеточное пространство пересекается мостиками
22
диаметром 7 нм на расстоянии до 10 нм. В некоторых случаях в мостиках отмечают наличие мельчайших пор. Соответствующие материалы получены и замораживанием — сколом. Это дает осно вание полагать, что глобулярная частица в области щелевого кон такта тянется через бислой липидов мембран и впячивается в межклеточную щель, где соединяется с соответствующей частицей противоположной мембраны смежной клетки. Соединение конец в конец этих частиц образует единицы — коннексоны, по которым из клетки в клетку идет гидрофильный канал диаметром 1,5—2нм, проводящий ионы и мелкие молекулы, поддерживая их электриче ские и метаболические взаимодействия. Проницаемость щелевых контактов достоверно доказывается прохождением при микро инъецировании флуоресцентных красителей, аминокислот, нуклеотидов и других веществ из одной клетки в другую. Белки, амино кислоты и другие макромолекулы через щелевой контакт не про ходят.
Рибосомы представляют собой гранулы 15—35 нм в диаметре. Располагаются они в цитоплазме свободно или фиксированы на мембране эндоплазматической сети (гранулярная эндоплазматическая сеть). Свободные рибосомы характерны для цитоплазмы недифференцированных камбиальных клеток. При световой мик роскопии цитоплазма клеток, богатых рибосомами, базофильна. Рибосомы имеются и в составе ядра, где они обеспечивают синтез ядерных белков (рис. 15).
Состоят рибосомы из двух субъединиц — малой и большой. Малая субъединица прикреплена к уплощенной области большой субъединицы. Каждая из них содержит молекулу рибосомальной РНК (р-РНК) и белка, который составляет 40—60% общей массы рибосомы. Располагаясь на мембранах эндоплазматической сети цитоплазмы клетки, рибосома прикрепляется большой субъеди ницей.
Рибосомы участвуют в сборке молекул белка — укладке амино кислот в полимерные цепи в строгом соответствии с генетической информацией, заключенной в ДНК.
Помимо рибосомальной РНК, в клетке присутствует информа ционная РНК (и-РНК), синтезирующаяся на ДНК ядра. Послед няя определяет порядок чередования азотистых оснований в п-РНК. и-РНК несет информацию от генома к рибосомам цито плазмы, где закодированное сообщение транслируется в последо вательность включения аминокислот синтезируемого белка.
Белок синтезируется обычно не на одной рибосоме, а на груп пе рибосом — полирибосоме (полисоме) . Рибосомы в полисоме связаны молекулой и-РНК, которая проходит вдоль ряда рибосом, пока вся закодированная в ней информация не будет прочитана. Информационная РНК связана с малой субъединицей рибосом, формирующаяся полипептидная цепочка — с большой.
Третий вид РНК в цитоплазме — это транспортная РНК (т-РНК), которая переносит аминокислоты на рибосому. Сущест вует специальная т-РНК для каждой аминокислоты, и каждая не-
23

Рис. 15. Полирибосомы ретикулоцитов: А — напыленные платиной (ув. 400 000); Б — окрашенные позитивно уронилацетатом (ув. 400 000, по Рич).
сет специфический тринуклеотид, способный прикрепляться к специфическому тринуклеотиду (кодону) па молекуле и-РНК. Последовательность ко донов на молекуле и-РНК определяет по следовательность прикрепления т-РНК и, следовательно, последо вательность чередования аминокислот в формирующейся полипептидной цепочке.
Рибосома создает пространственные отношения, необходимые для взаимодействия т~РНК с п-РНК; и обеспечивает формирование иолипептидных связей между аминокислотами, которое катализи руется активным участком одного из рибосомальных белков.
Эндоплазматическая сеть — система трубочек и уплощенных расширений, пазываемых цистернами, создающими в совокупности мембранную сеть в цитоплазме клетки. Эндоплазматическая сеть участвует в процессах синтеза, выполняет транспортную функцию в клетке, содержит ферменты и их субстраты, играющие активную роль в обмене веществ клетки. Различают два типа эндоплазматической сети: гранулярную, к наружной поверхности которой при креплены рибосомы, и агранулярную без рибосом (рис. 16).
Цистерны |
г р а н у л я р н о |
й |
э н д о п л а з м а т и ч е с к о й с е- |
т и особенно |
многочисленны |
в |
клетках, синтезирующих большое |
количество белка в качестве секреторного продукта. В таких клет ках цистерны могут располагаться параллельными скоплениями или образовывать концентрические системы. В таких скоплениях просвет цистерн очень узкий, расстояние между ними всего лишь
Рис. 16. Электронная микрофотография агранулярной (А) эндоплазматиче ской сети клеток печени хомяка (по Картези и Лонду) и гранулярной (В) эндоплазматической сети клетки поджелудочной железы.
25
/
1 — сигнальный кодон; 2 — и-РНК; г — сигнальный пептид; 4 — полость цистер-. ны; 5 — сигнальная пептидаза; б — рецепторный белок.
35 нм. На тангенциальных срезах цистерн гранулярной эндоплазматической сети видно, что рибосомы, фиксированные на ее мем бранах, также объединены в полисомы и расположены в виде ро зеток, спиралей, петель на внешней поверхности мембран.
Для объяснения прохождения синтезированных белков через мембрану в каналец эндоплазматической сети создана следующая гипотеза. Информационная РНК для секреторных белков содер жит последовательность сигнальных кодонов. Синтез сигнальных пептидов происходит на свободных рибосомах. Когда сигнальный пептид появляется из канала на большей субъединице, рибосома связывается с рецепторными белками для рибосом на мембране эндоплазматической сети. Такими белками являются, по-видимому, рибофорин I и рибофорин II, отсутствующие на мембранах глад кой эндоплазматической сети. Рецепторные белки при этом сбли жаются и образуется трансмембранный канал, расположенный так, что он является продолжением канальца на большой субъедипице рибосомы. В результате удлинения полипептидной цепочки сигнальный полипептид продвигается внутрь цистерны и отщеп ляется сигнальной пептидазой, локализованной на внутренней по верхности мембраны. Полипептидиая цепочка синтезирующегося секреторного белка продолжает продвигаться внутрь цистерны. Когда синтез белковой молекулы заканчивается, рибосома отделя ется от мембраны, а каналы облитерируются (рис. 16, а).
А г р а н у л |
я р и а я (гладкая) |
э н д о п л а з м а т и ч е с к а я |
сеть обычно |
не образует цистерн, а состоит из анастомозирую- |
|
щих трубочек. Она связана с синтезом и расщеплением гликогена,
с метаболизмом липидов, в частности, с синтезом стероидных гор монов. Поэтому гладкая эндоплазматическая сеть очень развита в клетках, продуцирующих стероидные гормоны (интерстициальных клетках семенника, клетках коры надпочечников, желтого тела яичников). Введение экспериментальным животным барбитуратов, инсектицидов, канцерогенов и других препаратов вызывает гипер трофию гладкой эндоплазматической сети в клетках печени. Этот адаптивный ответ печеночных клеток, повышающих свою способ ность метаболизировать и удалять лекарства, лежит в основе толе рантности к лекарствам при их продолжительном употреблении. Таким образом, агрануляриая эндоплазматическая сеть участвует в обезвреживающей функции печени.
Митохондрии присутствуют почти во всех эукариотических клетках. Их главная функция — обеспечение химической энергией, необходимой для биосинтетической и моторной активности кле ток. Продукты расщепления углеводов, поступающие в митохонд рию в виде пируватов, аминокислоты и жирные кислоты окисля ются в митохондриях до С02 и Н2 0. Освобождающаяся при этом энергия используется для синтеза аденозинтрифосфата (АТФ) из аденозиндифосфата (АДФ) и неорганического фосфата. Реа*[Ция формирования АТФ называется ф о с ф о р и л и р о в а и и ем, А1Ф обеспечивает энергией почти все жизненные процессы. При этом АТФ расщепляется на фосфат и АДФ. Последний вновь поглощает ся митохондрией и фосфорилизуется. Для обеспечения процессов окисления, фосфорилирования и других реакций в митохондриях
присутствует более 50 ферментов. На светооптическом уровне ми
тохондрии выглядят как нити или короткие палочки, реже — зерна. Их средняя длина 2—6 мкм, ши рина 0,2 мкм. Обычно они распре делены по всей цитоплазме, но иногда могут быть сконцентриро ваны на тех участках клетки, где потребность в энергии наибольшая. Например, вблизи аппарата движе ния (рис. 17).
Рис. 17. Митохондрии клеток приРис. 18. Схема общей организации мп-
.чматического эпителия кишечнитохондрии:
1 — внешняя мембрана; г — внутренняя мембрана; 3 — впячивание внутренней мем браны — кристы; 4 — места выпячиваний, вид с поверхности.
26

Рис. 19. Улектронная микрофотография среза митохондрии поджелудоч ной железы.
Митохондрии обладают характерной у л ь т р а с т р у к т у р о й . Снаружи митохондрия окружена гладкоконтурной наружной митохондриальной м е м б р а н о й толщиной 7 нм. На расстоянии 8—10 нм от наружной лежит внутренняя митохондриальная мем брана. Она имеет многочисленные складки — митохондриальные к рис ты, увеличивающие площадь внутренней мембраны. Между наружной и внутренней мембранами располагается мембранное пространство низкой электронной плотности. Пространство, огра ниченное внутренней митохондриальной мембраной, заполнено гомогенным или тонкозернистым м и т о х о н д р и а л ь н ы м мат- р и к с о м. В матриксе локализованы ферменты цикла трикарбоновых кислот, в котором пируваты, а также продукты расщепления белков и липидов окисляются до СОг и НгО. Внутренняя мембрана митохондрий содержит ферменты дыхательной цепи. На внутрен
ней |
мембране митохондрий |
располагаются м и т о х о н д р и а л ь |
н ы е |
с у б ъ е д и н и ц ы или |
э л е м е н т а р н ы е ч а с т и ц ы . Они |
представляют собой сферические частицы диаметром 9 нм, свя занные с мембраной стеблем шириной 3—4 нм и длиной 5 нм. В субъединицах имеются наборы ферментов, ответственные за фосфорилирование (рис. 18, 19, 20).
В |
матриксе митохондрий расположены м и т о х о н д р и а л ь |
ные |
г р а н у л ы . Они могут быть свободными или связанными с |
кристами. Размер их колеблется от 25 до 120 нм в зависимости от типа клеток и их функционального состояния. Высокая плотность обычно скрывает внутренню структуру гранул, но на тонких сре зах видно, что они разделены очень тонкими септами. Функция
гранул до сих пор окончательно не установлена, но полагают, что они являются местами связывания двухвалентных катионов, в ча стности Са++, и участвуют, таким образом, в поддержании посто янства содержания их в окружающей митохондрию гиалоплазме.
Митохондрии обладают своими собственными ДНК и РНК. На срезах митохондрий молекулы ДНК выглядят как ветвящиеся нити различной толщины, окруженные более прозрачным участком матрикса. Когда разрушенные митохондрии распределяются по по верхности воды, освобожденная ДНК выглядит, как нить толщиной 4 нм, длиной 5 мм, замкнутая в виде окружности. Циркулярная форма митохондриальной ДНК очень напоминает ДНК вирусов и бактерий. В митохондриальном матриксе присутствуют также частицы рибонуклеопротеида — рибосомы диаметром 10—15 нмг информационная и транспортная РНК, а также все необходимые ферменты для синтеза ДНК, РНК и белка. Однако из-за малой информации, заключенной в геноме митохондрий, они не могут синтезировать все свои компоненты и синтез большинства фермен тов обеспечивается геномом ядра.
Митохондрии обладают ограниченной продолжительностью су ществования (полупериод жизни для митохондрии клеток печени 8 дней, сердечной мышцы —6 дней, нейронов — 31 день). Убыльмитохондрий пополняется за счет их деления. При этом от внут ренней мембраны митохондрии растет септа, пока не встречается с противоположной стороной внутренней мембраны. В септу кон центрически врастает наружная мембрана и происходит разделе ние митохондрии на две дочерние.
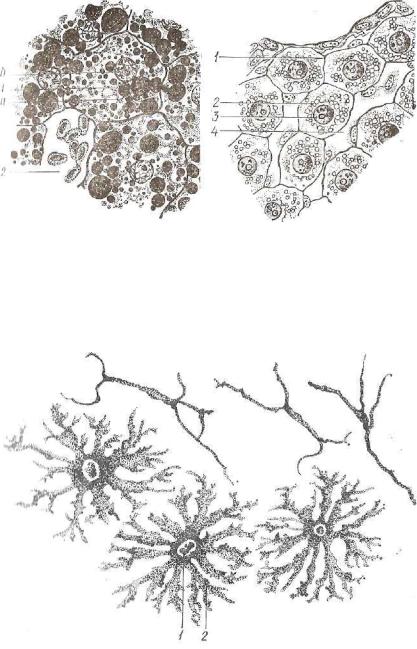
Комплекс Гольджи (пластинчатый комплекс) на препаратах, обработанных азотнокислым серебром или четырехокисью осмия, выглядит как сеть переплетающихся темных линий. В одних клет-
Рис 20 Электронная микрофотография кристы: митохондрии сер дечной мышцы быка (ув. 40 000). Видны частицы, переносящие электроны.
29
28

Рис. 21. Комплекс Гольджи (3) в нервных клетках спинального ганглия. Импрегнация осмием (ув. 400, по Алмазову и Сутулову):
1 — ядро с ядрышком; 2 — цитоплазма.
ках он локализован вблизи центриолей, в других — окружает ядро, а в эпителиальных клетках обычно располагается между ядром и апикальной поверхностью клетки (рис. 21).
При электронной микроскопии видно, что основным компонен том комплекса Гольджи являются окруженные мембраной упло щенные мешочки, или ц и с т е р н ы , располагающиеся стопкой друг над другом. Цистерны изогнуты так, что в стопке можно раз личить выпуклую (наружную) и вогнутую (внутреннюю) поверх ности (рис. 22). Отдельное скопление цистерн называется диктиосомой. Особенности ультраструктуры комплекса Гольджи связаны с его основной функцией конденсации и выведения секретов. Бел ковые секреты синтезирутся на рибосомах, связанных с грануляр ной эндоплазматической сетью, поступают в канальца сети и транспортируются в зону Гольдя-си. Цистерны эндоплазматической сети на поверхности, обращенной к комплексу Гольджи, обычно лишены рибосом. Маленькие выпячивания этой поверхности, за полненные белковым секретом, отрываются и образуют т р а н с п о р т н ы е пузырьки, вливающиеся в наружные цистерны диктиосомы. Помимо цистерн и транспортных пузырьков, в состав комплекса Гольджи входят также к о н д е н с и р у ю щ и е вакуо ли и с е к р е т о р н ы е г р а н у л ы . Согласно наиболее общепри нятой в настоящее время концепции, участие комплекса Гольджи в процессе секреции заключается в следующем. Транспортные пу зырьки, сливаясь, образуют цистерны наружной (формирую щей) поверхности диктиосомы. По мере формирования новых цистерн старые отодвигаются к внутренней ( с о з р е в а ю щ е й )
поверхности. Цистерны раздуваются, превращаясь в конденсирую щие вакуоли. Последние в результате конденсации их содержимого могут превращаться в секреторные гранулы. Мембраны цистерн по мере продвижения к созревающей поверхности трансформиру ются, приобретая сходство с плазмолеммой. Поэтому оболочка сек реторных гранул легко сливается с плазмолеммой и секрет посту пает в просвет железы. Лизосомы формируются в комплексе Гольджи так же, как секреторные гранулы (рис. 23).
Однако есть другая гипотеза, получившая в настоящее время довольно широкое распространение. По мнению ее сторонников, трубочки и цистерны с гладкой поверхностью, расположенные вблизи созревающей поверхности комплекса Гольджи и дающие реакцию на кислую фосфатазу, представляют собой специализи рованную систему для передачи кислых гидролаз из гранулярной эндоплазматической сети непосредственно в лизосомы, минуя комплекс Гольджи. Эта система была названа ГЭРЛ (связанный с комплексом Гольджи эндоплазматический ретикулум, от которого формируются лизосомы). Авторы связывают с ГЭРЛ также фор мирование пероксисом, конденсирующих вакуолей и мембран аутофагосом.
Помимо выведения белковых секретов, комплекс Гольджи при нимает участие в синтезе полисахаридов и присоединения их к белку. При синтезе гликопротеинов часть олигосахаридов вклю чается в полипептиды, когда они синтезируются на рибосомах, а. другие добавляются позднее к сформированным полипептидным цепям при достижении комплекса Гольджи. Кроме участия в син-
Рпс. 22. Электронная микрофотография комплекса Гольджи (стрелками обозначены мелкие вакуоли).
30 |
31 |
|
тезе углеводной части секреторных гликопротеинов железистых клеток, комплекс Гольджи играет важную роль в синтезе глико протеинов плазмолеммы (гликокаликса).
Морфология комплекса Гольджи может зависеть от интенсив ности процесса секреции. Когда органелла относительно неактив на, цистерны непрерывны, тесно расположены и одинаковой шири ны по всей диктиосоме. В активно секретирующих клетках профили цистерн короче, их мембраны часто фенестированы, а ширина про света увеличивается от формирующей поверхности к созревающей. В области комплекса Гольджи, кроме гладких, встречаются также •окаймленные пузырьки. В секреторных клетках их связывают с рециркуляцией мембран от плазмолеммы обратно к комплексу Гольджи.
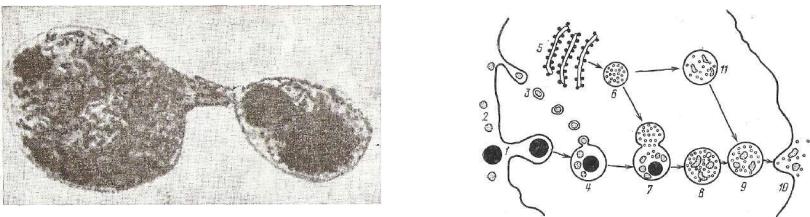
Лизосомы — тельца диаметром 0,2—0,5 мкм, ограниченные мембраной и содержащие около 50 различных ферментов, преиму щественно гидролитических, активных при кислых значениях рН (фосфатазы, гликозидазы, протеазы, липазы, сульфатазы и др.). Свое название органелла получила за то, что заключенные в ней ферменты способны вызвать лизис (растворение) всех компонен тов клетки. В нормальных условиях этого обычно не происходит, так как заключенные в лизосомах ферменты изолированы от суб стратов и соответственно неактивны. Около 20% ферментов встроено в мембрану лизосом и 80% находится в ее мукополисахаридном комплексе (рис. 24).
Функция лизосом заключается во внутриклеточном фермента тивном расщеплении как экзогенных веществ, попавших в клетку в результате эндоцитоза, так и эндогенных (удаление органелл и включений в ходе нормального обновления или в ответ на изменен ную функциональную активность). Иногда может повышаться проницаемость мембран лизосом клетки и их ферменты выходят
Рис. 24. Схема функционирования лизосом и внутриклеточного протеолиза (по Де-Дюву):
1 — фагоцитируемая частица; 2 — микромолекулы; 3 — макромолекулы, пиноцитируемые клеткой* 4 — фагосома; б — эргастоплазма; 6 — лизосомы; 7 — слия ние лигосомы и фагосомы; 8 — протеолиз частиц и макромолекул (9); 10 — экс креция остатков протеолиза; и — протеолиэ в лизосоме с образованием фагоцитоз* ной вакуоли.
в цитоплазму. Тогда происходит растворение (аутолиз) клетки. Это наблюдается в условиях эксперимента, патологии и в некото рых случаях нормального функционирования органа (инволюция молочной железы при прекращении лактации, инволюция матки после родов, резорбция хвоста амфибий при метаморфозе и др.). В зависимости от активности лизосом в процессах внутриклеточ ного переваривания и от характера объекта, подлежащего гидро литическому расщеплению, содержимое лизосом очень разнород но. Различают: первичные лизосомы, фаголизосомы (или гетерофагосомы), аутофагосомы и остаточные, или резидуальные, тельца.
П е р в и ч н ы е л и з о с о м ы — маленькие тельца с гомогенным содержимым. Они представляют собой резерв гидролитических ферментов, еще не участвующих в переваривании.
Ф а г о л и з о с о м ы (гетерофагические вакуоли) образуются от слияния первичной лизосомы с фагосомой. При этом начинается гидролитическое расщепление содержимого последней.
А у т о ф а г о с о м ы возникают при внутриклеточном обновле нии или при внутренней перестройке клетки, связанной с умень шением физиологической активности. Тогда часть органелл уда ляется путем аутофагии. Подлежащие разрушению органеллы окружаются мембраной, формируя аутофагическую вакуоль. С по следней сливаются лизосомы, изливая свои гидролитические фер менты. Природа мембраны окончательно не выяснена. По-видимо
му, это мембраны гладкой эндоплазматической |
сети, |
или ГЭРЛ. |
По мере переваривания содержимого фаголизосомы уменьша |
||
ются в размере Ж превращаются в о с т а т о ч н ы е , |
или р е з и |
|
д у а л ь н ы е , т е л ь ц а , заполненные гранулами |
непереваримого |
|
32
материала различного размера и плотности. Остаточные тельца впоследствии могут сливаться в скоплении липофусцина или пиг мента изнашивания.
Псроксисомы — окруженные мембраной сферические тельца размером 0,2—0,5 мкм. Они несколько напоминают лизосомы, но не содержат гидролитических энзимов. Для пероксисом характер но присутствие в них оксидаз аминокислот и каталазы, фермента, разрушающего перокиси. У тех видов животных, клетки которых содержат уратоксидазу, в пероксисомах печени и почек присутст вует кристаллоид (нуклеоид). У видов, лишенных уратоксидазы (птицы, человек), кристаллоид в пероксисомах отсутствует. Строе ние кристаллоида имеет видовые различия, а также зависит от ти па клетки. Например, в пероксисомах печени крыс кристаллоид состоит из полых трубок, расположенных таким образом, что они образуют фигуру пчелиных сотов. В пероксисомах проксимального отдела почек крысы обнаружены нуклеоиды двух типов: цилин дрические включения диаметром 85—140 нм и тубулярные кри сталлы длиной до 3 мкм. В пероксисомах печени хомячка нуклеоид, имеет форму пластинки.
В дополнение к пероксисомам печени и почек в различных ви дах эпителия обнаружены ограниченные мембраной тельца диа метром 0,15—0,25 мкм, лишенные нуклеоида —микропероксисомьь
Каталаза пероксисом может играть защитную роль, разрушая» перекись водорода, токсичную для клеток. Пероксисомы связыва ют также с метаболизмом холестерина, так как они особенно мно гочисленны в клетках, участвующих в метаболизме холестерина и синтезе стероидов: печени, надпочечников, яичников и в интерстициальных клетках семенников. Отмечено также, что введение ве ществ, понижающих уровень холестерина в крови, вызывает рез кое увеличение числа пероксисом печени.
Центросома (клеточный центр) располоясена вблизи ядра и комплекса Гольджи. На светооптическом уровпе она представлена двумя гранулами — ц е н т р н о л я м и, окруженными светлой бес структурной зоной цитоплазмы — ц е н т р о с ф е р о й . Последняя переходит в лучистую сферу, то есть в зону радиально расходя щихся тончайших, находящихся на грани микроскопического ви дения фибрилл (рис. 25).
При электронной микроскопии центриоли видны в виде цилин дров 300—500 нм длины и 150 нм в диаметре, стенка которых обра зована девятью группами микротрубочек. Каждая группа содержит 3 микротрубочки по 25 нм в диаметре. В группе микротрубочки располагаются цепочкой, ориентированной к радиусу центриоли под углом 40°. Микротрубочка, наиболее удаленная от периферии структуры, обозначается как субъединица А, остальные две, соот ветственно их положению, В и С. Микротрубочка А состоит из 13 тубулиновых протофиламентов. Микротрубочки В и С— из 10-11.
Перед делением клетки происходит удвоение центриолей, прш этом предсуществующие центриоли не делятся. Дочерняя цент-
Рис. 2П. Клеточный центр эпителиальной клетки: А, В, С — микротру- (И1чки центриоли.
|titu,iii. формируется заново па специфической области предсущест* iyiuii(oil центриоли, но будучи отделенной от нее узким пространОТИом, Вначале па боковой поверхности старой центриоли форми руется кольцевидное скопление плотного материала такого же диимотри, как зрелая центриоль, но лишенная микротрубочек — II р о ц о и т р и о л ь. Плотный материал продолжает присоединять ся к е.иободпому краю процентриоли, а в гомогенном, ранее плотМнм миторипло появляются триплеты микротрубочек. Формирую щимся цгптриоль удлиняется в паправлепии, перпендикулярном к мпторшюкой центриоли.
Функция цоитриолой заключается в индукции полимеризации Пилион - тубулиион с образованием микротрубочек. В интерфазе ОНИ У чисти у ют в формировании микротрубочек клеточного карка ли, III» прими митоза центриоли индуцируют формирование микро- •гругмишк перетони деления. Центриоли служат базальными тель-
ЦММИ |НШ1114 И К.
0 центриолью могут быть связаны сателлиты, представляющие еппом грииулнрпыо фокусы отхождения микротрубочек, и дополнlimnI.ими микротрубочки.
Н процессе формирования ресничек центриоли (базальное тельЦи) могут формироваться и на расстоянии от существующих ЦВНТриолий. большинство из них развивается вокруг плотных сфе рических тслоц, называемых д е у т е р о с о м а м и , или оргапизатоjiiiMii ироцоптриолей, которые, в свою очередь, развиваются путем
Л* |
35 |
1*110. 26. Гликоген в клетках печени. |
Рис. 27. Жировые включения в клет- |
|||
окраока кармином по методу Беста |
ках печени. Окраска осмиевой кис- |
|||
(у |
п. |
000): |
лотой — сафранином (ув. 900): |
|
I — плетки печени; 2 — цитоплазма с |
1 — клетки печени |
(а — липоидные гра* |
||
порнами и глыбками гликогена; з — яд- |
нулы в цитоплазме; |
б — ядро); 2 —ка-» |
||
lid о ядрышком; 4 — синусоидный (рас- |
пилляр с эритроцитами, |
|||
ширенный) кровеносный капилляр. |
|
|
||
PliO. 28. Пигментные включения в меланоцитах. Тотальный неокрашенный препарат (ув. 400):
/ - ядро пигментной клетки; |
2 — цитоплазма с пигментными зернами — мела* |
||
нином. |
|
|
^& |
диаметром 10 нм. Состоят они из белковых субъединяц. Белки в различных тканях отличны. В эпителии это кератины. Пучки мик рофибрилл в эпителии называются тонофибриллами. В клетках мезенхимного происхождения (фибробласты) микрофибриллы со стоят из белка виметина, в мышцах — десмина и скелетпна.
М и к р о ф и л а м е н т ы толщиной 6 нм в большом количестве присутствуют в кортикальном слое клеток, формируют пучки в их цитоплазме. Состоят они из сократительных белков, главным обра зом актина. В цитоплазме кровяных клеток гранулоцитов, фибробластов, нейронов и других клеток обнаружен также миозин.
Клеточные включения. В цитоплазме клеток различных тканей и органов в соответствии со специфичностью обмена веществ за кономерно синтезируются и накапливаются различные вещества в виде характерных для них продуктов обмена — включений. Они бывают трофические — связанные с белковым, углеводным и жи ровым обменами, секреторные, пигментные, включения витаминов
идр. Не являясь постоянной составной частью цитоплазмы, вклю чения отражают закономерности обмена соответствующих тканей
иорганов. Так, для яйцевых клеток характерны включения белка определенной химической характеристики. Накопление включений гликогена клетками печени соответствует закономерностям процес са пищеварения (рис. 26). Жировые включения физиологически накапливаются в жировых клетках соединительной ткани (рис. 27). Пигментные клетки эпидермиса кожи содержат включения мела нина (рис. 28). В клетках различных органов накапливаются ви тамины и многое другое.
ДЕЛЕНИЕ КЛЕТОК
Все клетки многоклеточного организма образуются последователь ным делением оплодотворенной яйцевой клетки. В процессе онто генеза клетки, размножаясь и дифференцируясь, сохраняя одина ковую генетическую информацию, приобретают характерные мор фологические особенности, специфические для различных тканей и органов данного вида организмов. Различают два вида клеточно го деления: митоз (греч. mitos — нить) и амитоз. Период, предше ствующий митотическому делению клетки, называется интерфазой.
Интерфаза характеризуется интенсивным ростом клетки и ее функциональной и морфологической специфичностью. По харак теру обмена веществ она слагается из трех периодов. В первый, пресинтетический, период (период Gi) клетка резко увеличивает ся в объеме. В ее цитоплазме активизируются процессы синтеза РНК, ферментов и других веществ, характерных для данного вида клетки. Второй период — синтетический (период S) —время удво ения молекул ДНК и синтеза белка гистона. В этом периоде к каждой хромосоме, содержащей после предыдущего деления одну хроматиду, достраивается парная хроматида. Процесс синтеза ДНК и включение в формирующуюся хроматиду белка гистона в отдель ных хромосомах клетки протекает последовательно в течение 5—6
38
и более часов. Последними включаются в процесс синтеза ДНК no il] сшие хромосомы.
Постсинтетическая фаза (третий период G2) значительно корочо диух предыдущих. В этот период синтезируются РНК и белки, участвующие в процессах деления клетки. В частности, синтези руются белки, входящие в состав структур цитоплазмы клетки, обеспечивающие процесс последующего митотического деления (тубулины ахроматинового веретена деления и др.). В течение промитотической фазы центриоли клеточного центра удваиваются. Oico.no каждой центриоли под прямым углом формируется дочер пни центриоль, соответственно клеточный центр в этом периоде иптерфазы состоит из двух групп центриолей.
Митоз. Митоз протекает в четыре фазы: профаза, метафаза, «инфаза и телофаза (рис. 29).
13 период профазы клетка выключается из специфической для иго функции и соответственно утрачивает связанные с этим ха рактерные специальные структуры (десмосомы, тонофибриллы, роснички и др.). В цитоплазме активизируется клеточный центр,
.что проявляется в последовательном расхождении сформирован ных и период профазы пар центриолей. Вокруг каждой пары — дочерних центросом — формируются радиально ориентированные м и к р отру бочки, образуя ее лучистую зону. Система микротрубочек между расходящимися дочерними клеточными центрами оформлнотой в «митотическое веретено». В результате выше описанных процессов в цитоплазме делящихся клеток в период профазы обрннуотся характерная для нее «ахроматиновая фигура» из двух днойиых групп центриолей, окружающих их звезд — радиально расходящихся микротрубочек — и «митотического ахроматинового норотеиа» — пучка микротрубочек между формирующимися дочер ними клеточными центрами (рис. 30, а, б, в).
Параллельно с динамикой структурной организации цитоплаз мы клетки в период профазы наблюдаются закономерные измене нии но одра. В частности, по мере вступления клетки в процесс митотического деления хромосомы инактивируются и соответстNtilliio опирализуются. В результате в ядре увеличиваются количевТйо и. размеры глыбок хроматина, которые, объединяясь, в сово купности образуют плотный клубок нитей. В связи с прекраще нном активности ядрышковых организаторов хромосом исчезают ними ндрышки. Период профазы завершается распадом мембраны ндорипн оболочки на сегменты, смешивающиеся с мембранами энДОШШИМйТИчоокоп сети. Хромосомы рассредоточиваются в цитонлниме (о),
Н хромосомах, содержащих две хроматиды, в области первич ной норетнжки иыниляотся центромера (кинетохор), связывающая хромосому по мере ее перемещения в экваториальную плоскость
Омикротрупочкими ахроматинового веретена в клетке.
Впериод м от а фа и ы митотический аппарат клетки завершает оное раивитив. Он состоит из центральных микротрубочек, распо ложен пых между цептриолями двух полюсов, митотического вере-
39