

Полигибридное скрещивание
Положение о независимом наследовании разных пар аллелей и признаков было подтверждено Менделем при изучении наследования трех пар признаков у гороха. Он скрещивал сорт растения с круглыми семенами (А), желтыми семядолями (В) и серо-коричневой кожурой семян (С) с сортом, форма семян которого морщинистая (а), семядоли зеленые (Ь), семенная кожура белая (с). Материнское растение имело генотип ААВВСС, отцовское — ааЪЪсс. Поскольку родители гомозиготны по всем трем парам признаков, у них образуется по одному типу гамет: ABC и аЪс. Гибриды Fi будут иметь генотип АаВЬСс (тригетерозигота). При
42
тригетерозиготности три пары разных аллелей находятся в трех разных парах гомологичных хромосом. В результате независимого сочетания хромосом (значит, и аллелей) из разных пар у гибрида Fi образуется восемь сортов гамет: ABC, ABc, АЪС, аВС, Abe, aBc, abc, аЪс. При самоопылении в результате случайного сочетания гамет в F2 получается 64 комбинации, включающие 8 фенотипов.
Мендель установил, что расщепление по фенотипу при три-гибридном скрещивании представляет собой сочетание трех независимых моногибридных расщеплений. Чем больше признаков, по которым отличаются взятые для скрещивания особи, тем сложнее расщепление и сильнее возрастает комбинативная изменчивость. Число возможных комбинаций гамет и число классов по фенотипу можно определить, пользуясь таблицей 3.
3. Количество фенотипов и генотипов в f2 при скрещивании родителей,
отличающихся разным числом пар признаков
(при полном доминировании)
Число возможных комбинаций
гамет

Число классов в F2
Число гамет,
образующихся
в F2
Число пар признаков
Пропорция
по фенотипу
по генотипу
полных рецессивов в
F2
|
1 |
2'=2 |
4'=4 |
2'=2 |
з'=з |
1/4 |
|
2 |
22=4 |
42=1б |
22=4 |
32=9 |
1/16 |
|
3 |
23=8 |
43=64 |
23=8 |
33=27 |
1/64 |
|
4 |
24=16 |
44=256 |
24=16 |
34=81 |
1/256 |
|
л |
2" |
4я |
2я |
3я |
1/4" |
Для того чтобы понять, почему в пределах популяции каждого вида животных наблюдается такое большое разнообразие в типе телосложения, размерах, продуктивности и т. д., можно произвести простые расчеты при помощи формулы 2й. Цифра 2 показывает, что набор хромосом диплоидный, я — гаплоидное число хромосом у определенного вида животных. Если отец и мать гетерозиготны только по одной какой-то паре аллельных генов, расположенных в каждой паре хромосом, то при полном доминировании каждого из признаков число определяемых этими аллелями возможных различных фенотипов у их потомков будет: у крупного рогатого скота 230, или более миллиарда, у свиней 219, или более 500 тыс., и т. д. Но животные, очевидно, могут отличаться друг от друга и по большему числу пар аллелей. Поэтому потенциальные возможности комбинативной изменчивости огромны, и становится понятным, почему в природе не встречается абсолютно похожих особей, за исключением однояйцевых близнецов.
Огромной заслугой Менделя является то, что в процессе своей работы он не только установил закономерности наследова-
43
СТРУЧКОВИДНЫЙ
ггСС
РОЗОВИДНЫЙ
RRcc
©
ГАМЕТЫ
ГГ.."'
ОРЕХОВИДНЫЙ
RrCc
ГАМЕТЫ
F,
ОРЕХОВИДНЫЙ
RrCc
ОРЕХОВИДНЫЙ
RRCc
ОРЕХОВИДНЫЙ
RrCC
розовидный
Rrcc
РОЗОВИДНЫЙ
RRcc
ОРЕХОВИДНЫЙ
RRCc
ОРЕХОВИДНЫЙ
RrCc
СТРУЧКОВИДНЫЙ
ггСс
ОРЕХОВИДНЫЙ
RrCC
ОРЕХОВИДНЫЙ
RrCc
СТРУЧКОВИДНЫР
ггСС
РОЗОВИДНЫЙ
Rrcc
ОРЕХОВИДНЫЙ
RrCc
ЛИСТОВИДНЫЙ
СТРУЧКОВИДНЫМ
гСс![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
ния признаков, но и открыл основные принципы (законы) наследственности:
дискретной (генной) наследственной детерминации признаков.Этот принцип лежит в основе теории гена;
относительного постоянства наследственной единицы (гена);
аллелъного состояния гена (доминантность и рецессивность).
\1
ВЗАИМОДЕЙСТВИЕ НЕАЛЛЕЛЬНЫХ ГЕНОВ
Иногда на формирование признака влияют две или несколько пар неаллельных генов. Проявление признака в этом случае зависит от характера их взаимодействия в процессе развития организма. В первом поколении появляется новый признак, которого не было у исходных родительских форм, и соотношение фенотипов во втором поколении будет иным. Рассмотрим несколько примеров различных типов взаимодействия генов.
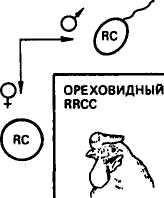
Новообразование. Новообразованием называется такой тип взаимодействия генов, когда при их сочетании в одном организме развивается совершенно новая форма признака. Известно,, что у кур гены розовидного и стручковидного гребня не являются аллельными. И стручковидный, и розовидный гребень доминирует над листовидным. При скрещивании кур породы виан* дот, имеющих розовидный гребень (RRcc), с петухами породы брама со стручковидным гребнем (rrCQ у потомков первого поколения (RrCc) в результате взаимодействия двух доминантных генов R и С развивается новая форма гребня — ореховидная (рис. 11). Скрещивание потомков Fi между собой ведет к получению в F2 четырех разных фенотипов в соотношении: 9 — с генами R и С с гребнем ореховидной формы, 3Rcc — с гребнем розовидной формы, ЪггС— со стручковидным гребнем и \rrcc <— с листовидным. Расщепление по фенотипу 9:3:3:1. В этом случае взаимодействие неаллельных генов R и С обусловливает образование новой формы гребня, в то время как каждый из этих генов в отдельности проявляет свой собственный эффект. Особь с листовидным гребнем является двойным рецессивом.
Комплементарное взаимодействие генов. В том случае, когда признак образуется при наличии двух доминантных неаллельных генов, каждый из которых не имеет самостоятельного фенотипи-ческого выражения, гены обозначают как комплементарные. Например, при скрещивании белых минорок с белыми шелковистыми курами первое поколение получается окрашенным. Для развития окраски необходимо, чтобы в организме синтезировались тирозин — предшественник меланина и фермент тирозингидрок-силаза, без которого пигмент не образуется. Обычно способность синтезировать какое-либо вещество доминирует над неспособностью к его образованию. Белые минорки имеют генотип ССоо. Они способны синтезировать тирозин, необходимый для образования
44
![]()
![]()
![]()
![]()
![]()
©
Рис. 11. Наследование формы гребня у кур при взаимодействии двух пар генов