

Основы альгологии и микологии Охапкин
.pdfdophyceae, Trentepohliophyceae). Основное цитологическое отличие классов – взаиморасположение базальных тел жгутиков и жгутиковых корней: при рассматривании подвижных клеток представителей этих классов сверху наблюдается сдвиг базальных тел относительно друг друга по часовой стрелке или против. У Chlorophyceae они сдвинуты по часовой стрелке как 1–7 часов или 12–6 на циферблате, у Ulvophyceae (в широком смысле) как 11–5 часов на циферблате, против часовой стрелки.
Ulvophyceae (в широком смысле) включает много морских обитателей и следующие порядки: Codiolales (Ulotrichales) (коккоидные, сарциноидные и нитчатые талломы), Ulvales (пластинчатые и трубчатые или мешковидные слоевища), Siphonocladales (талломы образованы многоядерными сегментами), Bryopsidales и Dasycladales (сложно устроенные талломы крупных размеров сифонового строения). Имеется смена форм развития, у большинства мейоз спорический.
Порядок Trentepohliales часто выделяют в самостоятельный класс Trentepohliophyceae с неопределенным систематическим положением. Подвижные клетки имеют верхушечные, а не боковые, как у харовых, жгутики; их базальные тела сдвинуты против часовой стрелки относительно друг друга, как 11 и 5 часов на циферблате, смена генераций изоморфная (как у Ulvophyceae). Жгутиковые корни в виде двух широких лент, цитокинез происходит с образованием фрагмопласта (как у харовых). Представители с гетеротрихальным талломом, ведут аэрофитный образ жизни.
Chlorophyceae включает водоросли монадной, коккоидной, нитчатой,
гетеротрихальной структур: порядки Volvocales, Tetrasporales, Chlorococcales, Chaetophorales, Oedogoniales. Исходными в морфологическом отношении являются одноклеточные монадные представители типа
Chlamydomonas из Volvocales.
Исходными коккоидными были одноклеточные водоросли типа Chorococcum из Chlorococcales. Нитчатая структура могла возникнуть и из монадной, и из коккоидной структур. С усложнением нитчатого строения сформировались гетеротрихальная (порядок Chaetophorales) структура. Оогамный половой процесс и своеобразное деление клеток привело к появлению талломов, характерных для представлителей порядка Oedogoniales. Преобладают гаплонты с покоящейся диплоидной зиготой, мейоз зиготический.
Зеленые стрептофитовые (харовые) (подотдел Streptophytina – отдел Charophya) включает несколько классов, наиболее крупные из которых
301
Zygnematophyceae и Charophyceae. Талломы одноклеточные, малокле-
точные, паренхиматозные, гетеротрихальные и сложно рассеченные (харофитные). Жгутиковые клетки асимметричны: жгутики субапикальные и латеральные, расположены под прямым углом к оси клетки. Мейоз зиготический, вегетативная фаза гаплоидная.
Мезостигматофициевые (класс Mesostigmatophyceae) – исходная предковая группа, от которой берет начало эволюция высших растений.
Порядок Zygnematales характеризуется отсутствием жгутиковых стадий в цикле развития; половой процесс – конъюгация, цитокинез идет с образованием фрагмопласта. Однако многие альгологи рассматривают их по-прежнему в качестве самостоятельного класса Conjugatophyceae (Zygnematophyceae) отдела Chlorophyta.
Порядок Klebsormidiales, имеющий черты примитивного строения, включает водоросли с сарциноидными и с нитчатыми неветвящимися талломами; половой процесс – изогамия.
Порядок Coleochaetales объединяет водоросли с разнонитчатым и паренхиматозным дисковидным талломом. Клетки соединены плазмодесмами; половой процесс – оогамия, бесполое размножение зооспорами. Зигота сохраняет хлоропласты и обрастает кроющими вегетативными клетками, которые образуются только после оплодотворения.
Центральный род порядка – Coleochaete (эпифиты подводных предметов и высших водных растений) – считается близким по структуре таллома к возможному предку высших растений в эпоху их возникновения. Были найдены подобные ископаемые формы. Признаки, сближающие с высшими растениями: многослойная структура, клеточное деление с образованием фрагмопласта, наличие фотодыхательного фермента гликолатоксидазы в пероксисомах.
Порядок Charales объединяет сложно устроенные водоросли мутов- чато-осевого строения с верхушечным ростом. Размножение вегетативное и половое, бесполого нет. Оогоний многоклеточный, единственная жгутиковая стадия – спермий. Ооспора развивается в предросток.
Согласно сравнительно-морфологическому анализу эволюция типов морфологической структуры в пределах классов Chlorophyceae, Ulvophyceae и Charophyceae шла параллельно. Предковыми формами были одноклеточные жгутиковые, сходные с современными представителями класса Prasinophyceae, которые во многих более ранних системах относили к семейству Polyblepharidaceae порядка Volvocales. Клетки и жгутики этих монад покрыты чешуйками, цитокинез многих представителей происходит с образованием фрагмопласта. У Nephroselmis или Py-
302
ramimonas жгутики боковые, жгутиковые корни в виде ленты. Возможно, предки этих родов дали начало Charophyceae.
Ряд представителей (Tetraselmis) покрыты текой, сходной с покровом хламидомонад, и цитокинез с фикопластом. Базальные тела жгутиков находятся в одной плоскости (12–6 часов на циферблате). Возможно, от них произошли Chlorophyta. От общего предка рано разошлись две линии, дав начало двум классам – Chlorophyceae (конфигурация базальных тел 1–7 часов на циферблате) и Ulvophyceae (в широком смысле) (соответственно 11–5 часов). Анализ рибосомальных генов показал большое сходство высших растений и харовых водорослей и различие между Chlorophyta и Charophyta.
Филогенетическое сходство высших растений с Prasinophyta и Chlorophyta, их большое сходство в строении и биохимических особенностях клеток теперь считается общепризнанным. В ряде систем эти группы объединяются в царство Viridiplantae или Plantae. Для всех этих групп характерны общие пигменты хлорофиллы а и b, основные ксантофиллы, наличие крахмала в качестве основного продукта ассимиляции и целлюлозы в клеточных стенках, путь синтеза лизина через диаминопимелиновую кислоту, сходные формы митоза, расположение тилакоидов в хлоропластах, наличие лишь двух мембран вокруг пластид, уплощенные или пластинчатые митохондриальные кристы, отсутствие фаготрофии и другие признаки.
У более примитивных высших растений гаметы имеют передние парные лишенные мастигонем жгутики, как у большинства зеленых водорослей. Основные отличия сводятся к большей сложности морфологии высших растений, которые всегда многоклеточны, большей частью дифференцированы на ткани и развиваются из зародыша, в который превращается зигота.
Гаметангии, если они не редуцированы, и спорангии у высших растений многоклеточные; наблюдается правильное чередование гаметофита и спорофита с разной степенью редукции одного из них. Все усложнения в организации высших растений связаны с их приспособлением к наземному образу жизни. Поэтому имеет смысл включать все группы зеленых водорослей с высшими растениями в одну группу (Streptophyta) и не разделять их по разным царствам растений, как у А.Л. Тахтаджяна (см. стр. 15).
303

ирУ·ОВП˚ ˝‚УО˛ˆЛЛ Л ЩЛОУ„ВМЛЛ ‚У‰УрУТОВИ1
èроблемы эволюции и филогении водорослей являются одними из самых важных и интересных проблем общей биологии. Их развитие охватывает огромный промежуток времени – от возникновения первых прокариот до настоящего времени – и связано с такими важнейшими событиями в эволюции органического мира на Земле, как появление фотосинтеза и фотоавтотрофных прокариот, возникновение эукариотной клетки, становление митоза, сингамии, т.е. слияния половых клеток с образованием зиготы, мейоза, смены ядерных фаз и форм развития (гаплоидной и диплоидной) и адаптацией к наземному сущест-
вованию.
Считают, что возраст прокариотических организмов приближается к возрасту древнейших горных пород, т.е. приблизительно 3,8 млрд лет. Первые автотрофные прокариоты – синезеленые водоросли или цианобактерии – появились 3,8–2,6 млрд лет назад в раннем докембрии. В протерозое, 2,6–0,6 млрд лет назад, эта группа организмов уже была представлена очень широко, и из ископаемых остатков, датируемых этим периодом, известны представители большинства порядков синезеленых водорослей, сохранившихся до наших дней. Каким же образом трактуется в современной науке вопрос о происхождении прокариот и фотоавтотрофов?
Предполагают, что возраст планеты Земля составляет 4–6 млрд лет. Примерно в течение половины этого периода в атмосфере Земли кислород отсутствовал. Первые этапы органической эволюции происходили, по-видимому, 4,5–3,1 (по некоторым данным 3,5) млрд лет назад, в так называемый биогенный период истории Земли. Этот период характеризовался переходом от неживой органической материи к клеточной организации жизни. Первичная атмосфера Земли преимущественно состояла из водорода, метана, азота, углекислого газа, сернистого ангидрида, хлористого водорода и других компонентов, но в ней не было свободно-
1 При написании этого раздела использованы материалы д.б.н., проф. Н.И. Стрельниковой (СПбГУ), за что авторы выражают ей искреннюю признательность.
304
го кислорода. В период так называемой химической эволюции, предшествовавшей эволюции биологической, на Земле, в результате сложного комплекса физико-химических процессов, происходило накопление различных классов органических веществ. Считают, что их концентрация происходила в водной среде, где и сформировались условия для биологического этапа эволюции. По-видимому, первые прокариоты появились в богатой органическими молекулами водной среде, при наличии атмосферы, носящей восстановительный характер.
Считают, что с позиций функциональной организации жизни первыми появились организмы, способные к брожению и обладающие функциями анаэробного обмена, т.е. у них сформировались фруктозобисфосфатный и пентозофосфатный пути метаболизма. Следующий этап эволюции жизни мог быть связан с имеющимися в водной среде сульфатами и с приобретением живыми организмами способности к электронному транспорту и формированию протонного потенциала как энергетического источника для регенерации АТФ. На этом этапе эволюции, возможно, появились содержащие железо и никель производные тетрапиррола – порфирины как компоненты электронно-транспортной цепи, а также автотрофный способ ассимиляции углерода – путь ацетилСоА. Таким образом, возникли анаэробно дышащие бактерии.
Следующим этапом функциональной эволюции прокариот могло быть возникновение фотосистемы I или протонного насоса, приводимого в действие энергией света. При этом живые организмы научились синтезировать (или поглощать извне) магнийпорфирины или хлорофиллы. Появилась способность фиксировать углекислый газ в рибулезобисфосфатном цикле с использованием неорганических доноров электронов (водорода, сероводорода, серы). Таким образом, возможно, возникли первые фототрофные организмы – прообразы современных фототрофных пурпурных и зеленых бактерий.
В дальнейшем в процессе эволюции, по-видимому, сформировалась фотосистема II, появился процесс нециклического переноса электронов с использованием воды как донора электронов. Этот процесс сопровождается выделением свободного кислорода и, таким образом, по-видимому, сформировался оксигенный фотосинтез у синезеленых водорослей, или цианобактерий. С появлением оксигенного фотосинтеза начинается формирование кислородной атмосферы Земли.
305
В среде, обогащенной кислородом, для защиты от его окисляющего воздействия живые организмы приобрели способность синтеза терминальных оксидаз из цитохромов и использования свободного молекулярного кислорода в качестве акцептора электронов. Таким образом, мог сформироваться аэробный путь дыхания у бактерий. Такая последовательность эволюции функциональной организации прокариот является лишь примером возможных событий, протекавших в эпоху их становления.
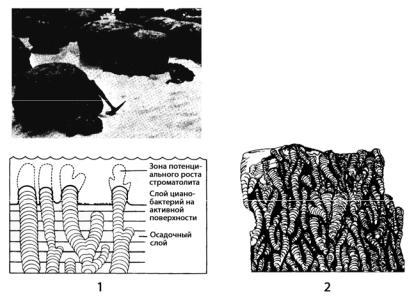
Предполагают, что первые прокариоты были водными анаэробными организмами. Их остатки в ископаемом состоянии встречаются в различных сланцах и строматолитах. Сланцы – это горные породы, которые характеризуются слоистым, ориентированным расположением породообразующих минералов и способностью раскалываться на тонкие пластинки. Различают сланцы кристаллические, например, слюдяные сланцы, гнейсы, и неметаморфизированные, например, глинистые, мергельные, углистые, битумозные. Строматолиты – это плоские, выпуклые, колончатые или куполовидные осадочные структуры с внутренней слоистостью (рис. 168).
Рис. 168. Строматолиты: 1 – внешний вид (сверху) и схема нарастания (снизу); 2 – продольный разрез
306
Считают, что они возникли как результат жизнедеятельности микроорганизмов и цианопрокариот, накапливающих в своих клетках или осаждающих различные химические соединения и образующих после отмирания микроорганизмов осадочные горные породы – строматолиты, возраст которых насчитвыает 2,7–3,5 млрд лет. Процесс образования строматолитов альго-бактериальными сообществами сохранился и до настоящего времени. В современных условиях строматолиты формируются в районах жаркого и сухого климата, в местах, где имеются мелкие водоемы.
Обилие строматолитов в ископаемых остатках свидетельствует о существовании аналогичных (или близких к ним) климатических условий в прошлые геологические эпохи, когда цианопрокариоты играли решающую роль в повышении уровня свободного кислорода в атмосфере ранней Земли. Если считать эти найденные остатки следами жизнедеятельности древнейших прокариот, то можно сделать вывод, что впервые земная жизнь должна возникнуть в промежутке между 3,5 и 4,6 млрд лет тому назад. Процесс образования строматолитов особенно интенсивно происходил в докембрии на протяжении позднего архея и протерозоя, т.е. примерно в течение 2,5 млрд лет, когда прокариоты были господствующей формой жизни на Земле.
Дискуссионным является вопрос, когда и в какой последовательности появились древнейшие прокариоты, поскольку и бактерии, и цианопрокариоты найдены в самых древних породах одновременно. Вопрос о происхождении первых оксигенных фотоавтотрофов – синезеленых водорослей – также проблематичен. Ряд авторов считает, что они возникли от эубактерий (так называемых настоящих бактерий), в частности, от анаэробных фотосинтезтрующих бактерий, представленных в современной микрофлоре тремя группами: зелеными и пурпурными серными и пурпурными несерными бактериями. Эти группы не используют воду в качестве донора водорода при фотосинтезе и выделяют не кислород, а ряд других продуктов, например, серу.
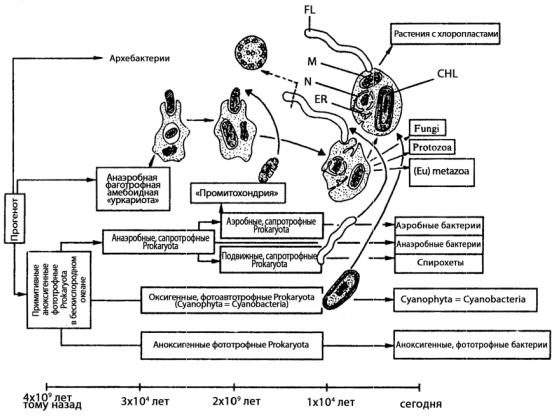
Рассматривают три древнейших эволюционных линии прокариот (эубактерии, уркариоты и архебактерии). Их выводят из общего предка, получившего название «прогенота» (рис. 169).
307
308
Рис. 169. Становление эукариотной клетки путем ассоциации нескольких различных прокариотных клеток с Ур-кариотой: CHL – хлоропласт; ER – эндоплазматический ретикулум; FL – жгутик (со структурой 9-9+2); M – митохондрия; N – ядро
Считают, что уркариоты представлены ядерно-цитоплазматическим компонентом эукариотной клетки, т.е. они, по-видимому, являлись примитивными анаэробными фаготрофными амебоидными организмами. В дальнейшем уркариоты включили в себя в качестве эндосимбионтов представителей различных групп эубактерий, превратившихся в митохондрии и хлоропласты. Эти три линии были выделены на основании анализа нуклеотидных последовательностей в молекулах рРНК (16S у прокариот и 18S у эукариот), состоящих из 1600 и 2500 нуклеотидов соответственно. Этот анализ выявил всех эукариот, кроме их органелл (митохондрии и хлоропласты), – это первая группа, представленная ядерно-цито- плазматическим комплексом эукариотной клетки. Ко второй группе, получившей название истинных бактерий, или эубактерий, относится большинство прокариот. В эту же группу попали на основании степени гомологии 16S рРНК митохондрии и хлоропласты эукариотных клеток. В третью группу вошли малоизученные, обитающие в экстремальных условиях прокариоты: метанобразующие бактерии, экстремальные галофилы и термоацидофилы, которым дали название архебактерий.
Анализ последовательностей нуклеотидов 16S рРНК у эубактерий показал, что фотосинтезирующие эубактерии не образуют единой родственной группы, а они были обнаружены в пяти группах бактерий из десяти выделенных. Это говорит о том, что фотосинтезирующие бактерии более родственны с представителями своей группы, чем с фотосинтезирующими видами из других групп. По-видимому, бактериальные фотосинтетики являются крайне древними, и многие нефотосинтезирующие группы бактерий возникли из фотосинтезирующих предков.
Также невозможно судить с уверенностью, использовали ли первые синезеленые водоросли обе фотосистемы – I и II, т.е. высвобождали ли они кислород как побочный продукт фотосинтеза. Поэтому сейчас нельзя с уверенностью говорить о происхождении цианопрокариот от предположительно фотосинтезировавших, но не высвобождавших кислород предков, подобных фотосинтезирующим бактериям.
Другие авторы считают, что накопленные к настоящему времени палеонтологические данные об остатках первых прокариот из архейских отложений недостаточны, чтобы с уверенностью отличить первых синезеленых водорослей от других прокариотических организмов. Эти находки однозначно указывают на облигатное существование цианобактериального автотрофно-гетеротрофного сообщества, которое осуществляло круговорот углерода на древней Земле и по своему происхождению было, вероятнее всего, полифилетичным. Устойчивое сущест-
309
вование таких сообществ на первых этапах эволюции биосферы в течение 2,5 млрд лет было возможным только при строгой сбалансированности по углероду продукционно-деструкционных процессов. Центром таких древних сообществ могли быть только цианопрокариоты, поскольку они являются основными продуцентами органического вещества. Поэтому высказывается мысль о независимости формирования в архее автотрофных и гетеротрофных путей метаболизма живых организмов.
Существуют весомые доказательства в пользу первичной автотрофности самых древних организмов. У них оксигенный фотосинтез мог сформироваться в локальных геохимических нишах в окислительных условиях среды. Возникновению оксигенного фотосинтеза могло способствовать наличие пероксида водорода, который в те времена изобильно образовывался из паров воды абиогенно под влиянием жесткого ультрафиолетового излучения, достигавшего поверхности Земли в условиях отсутствия озонового слоя. Как уже говорилось, в докембрийских бассейнах цианобактериальные маты занимали, вероятно, все подходящие местообитания – от мелководных участков суши до открытого моря. Это предположение позволяет объяснить поразительную быстроту начальных этаповэволюциижизнииисключить постулированиесуществованиявэтот период прародителя всех организмов – прогенота. По ряду цитологических и биологических признаков (например, размеру генома) и результатам секвенирования 16S рРНК синезеленые водоросли более сходны с оксигенными эукариотами. Поэтому нет достаточных оснований относить синезеленые к эубактериям и выводить их из эубактерий.
По косвенным признакам данных геологической летописи можно предположить, когда в архее начался кислородный фотосинтез
(рис. 170).
Сначала кислород выделялся в гидросферу и тратился на окисление восстановленных соединений, в конечном итоге попадавших в донные отложения. В период от 3,5 до 2,0 млрд лет назад на Земле происходило накопление и отложение геологических формаций, содержащих железо и слоистых по своей структуре. Эти отложения напоминают современные сланцы, слои которых то бедны, то богаты железом. В слоях, богатых железом, оно находится в трехвалентном окисленном состоянии. По-видимому, в окислении и осаждении в донные осадки двухвалентного железа, растворенного в морской воде, принимал участие кислород, высвобождаемый цианопрокариотами в процессе фотосинтеза. Таким образом, возможно, появление фотосинтеза произошло либо раньше, либо одновременно с появлением таких железистых полосчатых формаций (т.е. свыше 3,5 млрд лет назад).
310