

Основы альгологии и микологии Охапкин
.pdfотсутствию способности изменять форму тела, например, у видов рода факус (Phacus). В местах перекрытия белковых полос изнутри располагаются слизистые тельца, которые через узкий канал выделяют слизь наружу клетки. Выделение слизи происходит при образовании пальмеллевидных состояний и хорошо выражено у колониальных форм.
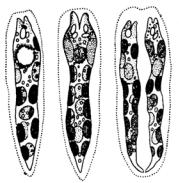
Клетки некоторых эвгленовых, например, у видов трахеломонас (Trachelomonas) (рис. 150) наружи окружены твердыми домиками шаровидной, яйцевидной, цилиндрической или веретеновидной формы.
Домики могут быть гладкими, либо украшены шипами, бугорками или различными другими выростами, бесцветные или окрашены солями железа в различные оттенки коричнево-бурых тонов. На переднем конце домик имеет отверстие, у многих видов окруженное горлышком. Стенки домика состоят из уплотненных слизистых веществ полисахаридной и белковой природы, пропитанных солями железа.
На переднем конце тела имеется глотка, состоящая из верхней, более узкой части – канала, и нижней, расширенной части – резервуара или ампулы, в полость которой изливается содержимое пульсирующих вакуолей (рис. 151).
Рис. 151. Схема ультратонкого строения жгутикового аппарата эвгленовых:
1– двигательный жгутик;
2– мастигонемы;
3– параксиальный тяж;
4– короткий жгутик;
5– жгутиковый резервуар;
6– кинетосома;
7– ленты микротрубочек;
8– парафлагеллярное тело; 9 – глобула стигмы;
10 – цитостом;
11 – промежуточный корешок из микротрубочек
У части голозойных видов глотка функционирует как цитостом, или клеточный рот, к ней примыкают органеллы, предназначенные для захвата пищи: так называемый палочковидный орган, или сифон. У фотосинтезирующих эвгленовых (эвглена – Euglena) недавно обнаружен рудиментарный цитостом, гомологичный цитостому бесцветных представителей отдела. За счет глоточного палочковидного аппарата хищные
271
гетеротрофные эвгленовые могут питаться простейшими, мелкими многоклеточными животными и другими объектами.
У фотосинтезирующих окрашенных видов есть хлоропласты, у гетеротрофных они отсутствуют. Число хлоропластов от одного – двух до нескольких сотен. Они имеют дисковидную, звездчатую или лентовидную форму, покрыты оболочкой из трех мембран. Крупные хлоропласты имеют пиреноиды, на поверхности которых откладывается особый запасной углевод эвгленовых – парамилон (полимер глюкозы). У одних видов парамилоновые зерна мелкие и заполняют всю цитоплазму, у других кроме мелких зерен формируются 1–4 крупных парамилии определенной формы (цилиндрические, дисковидные). У некоторых бесцветных эвгленовых обнаружены гликоген, жиры, гематохром (раствор астаксантина в масле), окрашивающий клетки в красный цвет (Euglena rubra). Тилакоиды в ламеллах группируются по три. Клетки имеют многочисленные митохондрии, одну сократительную вакуоль, формирующуюся из мелких вакуолей возле стенки жгутикового резервуара.
Ядро одно, находится в центральной, реже в задней половине особи, с одним или несколькими ядрышками. Для эвгленовых характерен митоз с сохранением ядерной оболочки с внутриядерным веретеном – эвгленомитоз. Ядрышко при митозе сохраняется и делится перешнуровкой. Хромосомы постоянно конденсированы.
Жгутиков 1–2, реже больше. У большинства видов жгутиков 2 не равных по длине и гетеродинамичных, только у немногих (например, у эвтрепции – Eutreptia, рис. 148: Д) оба жгутика почти одинаковые. Часто у двужгутиковых форм (Euglena) один жгутик сильно редуцирован, не выходит за пределы глотки и не виден в световой микроскоп. Наружная часть длинного жгутика покрыта тонкими мастигонемами или более сложно устроенным покровом из тонких волосков. Вдоль аксонемы (внутренняя система микротрубочек по схеме 9-9+2) проходит внутренняя опорная структура жгутика – параксиальный тяж. Он состоит из микрофиламентов и расположен между плазмалеммой и аксонемой, проходя по всей длине жгутика (рис. 151).
Фототрофные эвгленовые водоросли у основания длинного жгутика имеют расширение или вздутие – парафлагеллярное (парабазальное) тело (рис. 148: А; 151: 8), напротив которого вблизи дорсальной стенки жгутикового резервуара располагается стигма (глазок). Кроме окрашенных, стигма есть и у некоторых бесцветных представителей. Стигма состоит из отдельных шаровидных глобул, каждая из которых покрыта мембраной и заполнена липидами, в которых растворен β-каротин (рис.
272
14: 3). При затенении парафлагеллярного тела стигмой эвглена изменяет направление своего движения. Таким образом, стигма в совокупности с парафлагеллярным телом составляет две части фоточувствительного аппарата.
Эвгленовые водоросли могут плавать в толще воды за счет деятельности жгутика, у видов, обитающих на поверхности субстрата и ведущих ползающий образ жизни, наблюдается метаболия. У части видов жгутиковые и метаболические движения осуществляются одновременно. Часть видов вегетирует в пальмеллевидном состоянии, некоторые ведут прикрепленный образ жизни, лишь временно образуя монадные стадии (например, колациум – Colacium, рис. 148: Е).
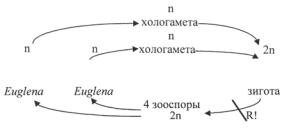
Размножение эвгленовых водорослей происходит путем продольного деления клетки надвое в подвижном или пальмеллевидном состоянии. Деление особей идет от переднего края клетки к заднему (рис. 24; 152), за исключением видов рода колациум (Colacium), ведущих прикрепленный образ жизни. У них этот процесс начинается с морфологически заднего свободного конца. У немногих видов описана хологамия и автогамия. Однако половой процесс у эвгленовых изучен недостаточно и считают, что он не является характерной чертой цикла развития.
Эвгленовые распространены по всему земному шару, являясь обитателями как морских (многие гетеротрофные эвглениды), так и пресных вод (как автотрофные, так и гетеротрофные представители). Наиболее многочисленны эвглено-
вые в мелководных стоячих пресных водоемах, где вода хорошо прогревается, содержит много биогенных и органических веществ. Большинство видов развивается в поверхностных или придонных слоях воды, некоторые ползают по дну. Немногие бесцветные виды обнаружены в почвах, представители рода Colacium развиваются как эпибионты на различных планктонных ракообразных. Паразитические формы эвгленовых найдены в кишечнике олигохет, пиявок, нематод, копепод, амфибий, на жабрах рыб. Отдел включает один класс и несколько порядков.
273
Класс Euglenophyceae – Эвгленовые – объединяет организмы с хлоропластами или без них, питающиеся фотоавтотрофно, миксотрофно или сапрофитно, редко питание голозойное. Некоторые виды являются паразитами или симбионтами животных.
Порядок Euglenales – Эвгленовые. Подвижные клетки всегда одиночные, но в пальмеллевидном состоянии образуют скопления, некоторые ведут прикрепленный образ жизни, образуя временную жгутиковую стадию. Отложения парамилона хорошо заметны в форме мелких или крупных зерен – парамилий. Питание фототрофное, миксотрофное или сапрофитное. У большинства представителей основной формой передвижения является плавание в толще воды, сопровождающееся вращением тела вокруг продольной оси. Есть виды, одновременно хорошо плавающие и ползающие или только ползающие по субстрату; ряд форм ведет неподвижный образ жизни. Наибольшим числом видов отличаются роды эвглена (Euglena),
трахеломонас(Trachelomonas) ифакус(Phacus).
Цикл развития Euglena
У эвглен клетки в разной степени метаболичные, вытянутые в длину, иногда в той или иной мере спирально скрученные (рис. 148: А; 152). Передний конец закруглен или вытянут, задний также закруглен или, чаще, сужен и заострен, у ряда видов заканчивается шиповидным отростком. Наружный жгут один, но со дна резервуара, которым кончается глотка, отходит другой, более короткий жгутик, не выходящий за пределы глотки и соединенный с длинным наружным. Хлоропласты зеленые, различной формы, от одного до многих, большей частью стенкоположные, реже осевые, с пиреноидами или без них. Имеется глазок или стигма. Ядро с ядрышком лежит обычно в средней части клетки. Питание преимущественно миксотрофное. Размножается делением. Известно пальмеллевидное состояние. Некоторые виды образуют цисты.
274
Виды рода Euglena обитают в малых и мелких водоемах, лужах, прибрежной полосе пресных, часто гумифицированных или слабо засоленных озер и рек, также морей, в болотах, на сыром грунте. Ряд видов, развиваясь
вмассевтеплоевремя года, можетвызывать «цветение» воды.
Утрахеломонас (Trachelomonas) (рис. 2: 5–8; 148: Б; 150) клетки ме-
таболичные, интенсивно плавающие, свободно располагаются внутри твердых, обычно бурых домиков, различной формы и строения. Стенки домиков гладкие или орнаментированные, с отверстием для выхода жгутика. Вокруг жгутикового отверстия стенки домика часто валикообразно утолщены, иногда отверстие окаймлено зубчиками или кольцевым выростом – воротничком или горлышком. Задний конец домика закруглен или сужен, иногда с длинным и узким полым отростком. Хлоропласты с пиреноидами или без них. Парамилон в виде мелких округлых, реже продолговатых зерен, иногда откладывается масло. Размножается делением клеток внутри домиков, после чего одна из особей выскальзывает из отверстия домика наружу и выделяет новый домик (рис. 24). Виды рода Trachelomonas широко распространены преимущественно в малых пресноводных водоемах, некоторые их них вызывают «цветение» воды.
Виды рода факус (Phacus) (рис. 3: 2, 3; 148: В) обитают в основном в небольших стоячих загрязненных органическими веществами водоемах, прибрежьях озер и рек, особенно заболоченных. Особи плавают свободно, неметаболичные, с плоско сжатым (до листовидного) телом, обычно заканчивающимся на заднем конце бесцветным прямым или изогнутым отростком. На одной из сторон клетки, которую считают спинной, расположена продольная складка в виде гребня. Клетки у большинства видов более или менее спирально скручены по продольной оси. Пелликула плотная, с продольными и косыми рядами штрихов, гранул, шипиков и др. Парамилон откладывается в виде 1–2 крупных парамилий и многих мелких зерен, рассеянных в цитоплазме.
Порядок Peranematales – Перанемовые – включает водоросли с 1– 2 большей частью неодинаковыми жгутиками; клетки неметаболичные или метаболичные, без хлорофилла и стигм. Клетки дорсовентральные, уплощенные, часто асимметричные. Жгутиковое отверстие функционирует как цитостом, имеется цитопрокт – аналог анального отверстия. У ряда видов есть палочковидная органелла, которая выворачивается наружу и прикрепляется к добыче; у других ту же функцию выполняет глоточный аппарат в виде трубки – сифон. Питание голозойное, реже
275
сапрофитное. Размножение продольным делением, у одного вида описана хологамия.
Перанема (Peranema) (рис. 148: Ж) обитает в стоячих и слабо проточных водах, в мелководной зоне у дна. Клетки сильно метаболичные, цилиндрические или веретеновидные с одним длинным, направленным вперед жгутиком. Второй – более короткий и тонкий жгутик тянется вдоль брюшной стороны клетки назад. К глотке примыкает палочковидная органелла. Питание голозойное и возможно сапрофитное.
У энтосифона (Entosiphon) (рис. 148: Г) клетки неметаболичные, овальные или яйцевидные, немного сжатые с твердой, продольно заштрихованной или ребристой пелликулой. Имеется два жгутика, из них передний, более короткий и тонкий, при движении интенсивно колеблется, а задний – более толстый и длинный, при движении волочится сзади. Рядом с глоткой находится узкоконическая органелла – сифон. Распространен в стоячих и слаботекучих водоемах.
Положение эвгленовых в системе эукариот трактуется неоднозначно. Наиболее примитивные, предковые формы, по-видимому, были схожи с современными бесцветными гетеротрофными эвгленовыми. Их эволюция, вероятно, шла в двух направлениях. Одна из групп в результате приобретения путем эндосимбиоза хлоропластов дала современных фототрофных и сапрофитных представителей порядка Euglenales. Другая группа превратилась в специализированных, питающихся преимущественно голозойно перанемовых – порядок Peranematales. Некоторые авторы считают, что современные бесцветные формы произошли от исходно зеленых. Скрытый цитостом зеленых эвгленид свидетельствует об их происхождении от бесцветных предков. Близкие филогенетические связи между одноклеточными животными кинетопластидами (бодониды) и бесцветными эвгленовыми доказывает сходство в строении их трихоцист. Данные анализа структуры рибосомальной РНК подтверждают филогенетическую близость эвгленовых и кинетопластид при их значительной удаленности от других эукариот. Наличие в хлоропластах некоторых эвглен белоксинтезирующих систем с чертами, свойственными прокариотам, и анализ последовательностей 16S рРНК и 28S рРНК свидетельствуют о древности ветви эвгленовые – кинетопластиды в системе эукариот. Другие молекулярно-биологические данные этого не подтверждают.
В современных системах протозоологов выделен тип Euglenozoa с двумя классами Euglenoidea и Kinetoplastidea на основании наличия скрытого цитостома у зеленых эвгленид, особенностей строения жгути-
276
ков, с присутствием сходных белков в параксиальных тяжах жгутиков, особенностей деления ядра. Все эти признаки могут служить доказательством филогенетических связей между обеими группами и подтверждают происхождение фотосинтезирующих эвгленовых от бесцветных гетеротрофных форм.
йЪ‰ВО Streptophyta – лЪрВФЪУЩЛЪУ‚˚В, ЛОЛ лЪрВФЪУЩЛЪ˚
ä этому отделу относят растения, клетки которых или не имеют жгутиков (Zygnematophyceae), или при их наличии жгутики за-
нимают боковое положение (Klebsormidiophyceae, Charophyceae). В
процессе цитокинеза так же, как у Trentepohliophyceae, новые клеточные стенки закладываются с участием фрагмопласта.
Подотдел Streptophytina – Зеленые водоросли стрептофиты.
Эта группа объединяет все Streptophyta, от одноклеточных до многоклеточных водорослей, имеющие талломную организацию. Наиболее сложно организованные водные формы достигают высокой функциональной дифференцировки, у них намечается тенденция к формированию тканей и защищенных органов размножения. По объему соответствуют отделу Charophyta, выделяемому некоторыми альгологами. Это линия преимущественно пресноводных зеленых водорослей, давшая начало высшим растениям. На родство с высшими растениями указывают сходные механизмы образования целлюлозы клеточной стенки, наличие многоклеточного таллома, фрагмопласт в цитокинезе, наличие плазмодесм, осуществляющих взаимодействие между клетками многоклеточного организма, наличие меристематических апикальных клеток (у харовых и колеохетовых), сложно ветвящиеся талломы, тенденция к образованию тканей, специфический фермент фотодыхания (гликолатоксидаза) и ряд других.
Водоросли класса Mesostigmatophyceae – Мезостигмофициевые –
это одноклеточные пресноводные монадные организмы (например, мезостигма – Mesostigma), покрытые покровом из трех слоев чешуек. Два немного неравных жгутика прикреплены не терминально. Хлорокибус (Chlorokybus) имеет сарциноидные талломы из округлых или эллипсоидных клеток, покрытых толстым слоем слизи. Строение зооспор хлорокибуса обнаруживает общие черты с организацией клетки мезостиг-
277
мы. В некоторых системах рассматривают отдельно два класса: Mesostigmatophyceae и Chlorokybophyceae. Принадлежность этой группы во-
дорослей к Streptophytina подтверждается анализом ДНК. По-видимому, мезостигмофициевые – это древняя исходная группа, от которой берет начало эволюция зеленых наземных растений.
Класс Zygnemophyceae (Conjugatophyceae) – Зигнемофициевые
(Конъюгаты) – это большая группа зеленых водорослей, в основном представленная пресноводными организмами. Сюда относятся одноклеточные, редко колониальные и многоклеточные водоросли, коккоидной и нитчатой структуры. Цитокинез идет с участием примитивного фрагмопласта. Характерной чертой этой группы является полное отсутствие жгутиковых стадий в жизненном цикле и наличие своеобразного полового процесса – конъюгации. Конъюгация заключается в слиянии недифференцированных протопластов двух вегетативных клеток. Между конъюгирующими клетками формируется конъюгационный канал; их протопласты сжимаются и перетекают навстречу друг другу, сливаясь в конъюгационном канале или внутри оболочки одной из родительских клеток. Зигота после периода покоя и редукционного деления прорастает с образованием 1, 2 или 4 проростков. Помимо конъюгации широко распространено вегетативное размножение делением клеток у одноклеточных и фрагментацией нитей; известны апланоспоры и акинеты. В разных системах у конъюгат выделяют от двух до четырех порядков.
Виды порядка Mesotaeniales – Мезотениевые – являются наибо-
лее примитивно устроенными представителями конъюгат. Клетки их организованы просто, не сегментированы, с гладкими, одноили двуслойными, всегда покрытыми обильной слизью, слегка инкрустированными известью оболочками. Индивиды одноклеточные эллипсоидной, веретеновидной или цилиндрической формы, преимущественно простые, реже изогнутые, без перетяжек посередине. Редко водоросли объединяются в бесформенные слизистые колонии или короткие нити. Оболочка клетки слоистая и гладкая, всегда покрыта обильной слизью. Хлоропласты лентовидные, спирально закрученные, или осевые пластинчатые с пиреноидами, или состоящие из центральной части и радиально отходящих от нее пластинчатых выростов, или длинных продольных ребер.
У рода мезотениум (Mesotaenium) (рис. 153: 1) осевой пластинчатый хлоропласт с одним или несколькими пиреноидами.
278
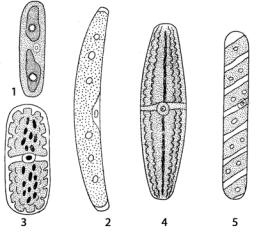
Рис. 153. Мезотениевые водоросли:
1 – Mesotaenium;
2 – Roya;
3 – Cylidrocystis;
4 – Netrium;
5 – Spirotaenia
У ройи (Roya) (рис. 153: 2) клетки цилиндрические, изогнутые, с зауженными и закругленными концами. На пластинчатом хлоропласте появляются продольные складки и срединный вырез у вогнутой стороны клетки, в котором помещается ядро. В центре клетки цилиндроцистиса (Cylindrocystis) (рис. 153: 3) находится ядро, по обе стороны которого симметрично располагаются два хлоропласта, каждый из которых состоит из центральной части и отходящих от нее небольших радиально расположенных пластин. У нетриума (Netrium) (рис. 153: 4) в клетке расположены два центральных хлоропласта, в поперечном разрезе имеющие звездчатую форму и состоящие из центральной осевой части и длинных ребер с изрезанными краями, расходящимися от нее по радиусам. У спиротении (Spirotaenia) (рис. 153: 5) одиночные клетки внешне похожи на клетки одноклеточных видов спирогиры (Spirogyra). Она имеет спирально закрученный пристенный хлоропласт или спирально закрученные ребра вокруг осевой части хлоропласта. Пиреноиды крупные, расположены в центре хлоропласта по 1–2 и более, реже они беспорядочно расположены по всему хлоропласту. Размножение вегетативное путем деления клеток в поперечной плоскости.
При конъюгации две клетки сближаются, выделяют обильную слизь, располагаются длинными осями либо параллельно, либо перпендикулярно друг другу. В месте контакта клетки вырабатывают отростки, ко-
279
торые при разрастании несколько отодвигают партнеров. В месте соединения отростков в их оболочках образуется отверстие и формируется конъюгационный канал. Протопласты клеток отстают от оболочек, сжимаются, перетекают навстречу друг другу через конъюгационный канал и там сливаются с образованием зиготы. Диплоидная зигота одевается многослойной оболочкой, превращаясь в зигоспору. Иногда перед конъюгацией содержимое клеток делится на две гаметы, а после их слияния формируются две зигоспоры. После периода покоя зигоспора прорастает мейотически, давая начало четырем новым дочерним гаплоидным организмам. Реже зигоспора прорастает двумя проростками. Мезотениевые – это небольшая группа, объединяющая 40–50 видов водрослей, распространенных в торфяных болотах, реже на сырой земле в виде слизистых скоплений.
К порядку Zygnematales – Зигнемовые – относятся нитчатые неветвящиеся водоросли, состоящие из одного ряда клеток, плотно примыкающих друг к другу. Размеры нитей у некоторых видов могут достигать 10 см. Оболочка клеток цельная, двуслойная, снаружи покрытая сильно развитым слизистым чехлом. Рост нитей в длину осуществляется за счет поперечного деления клеток. Наличие обильной слизи и яркозеленая окраска нитей, иногда с желтоватым оттенком, хорошо отличают зигнемовые нитчатые водоросли от представителей других порядков с нитчатой структурой таллома. Нити лежат на дне водоема почти свободно, или прикрепляясь к субстрату и образуя сплошной покров, или же плавают у поверхности воды на мелководье водоемов разного типа (прудов, озер, заводей рек) в виде «тины». У некоторых видов известны ризоиды – бесцветные выросты клетки с более толстой оболочкой, лишенные хлоропластов. Ризоиды чаще развиваются в условиях более высокой гидродинамики как средство выживания водоросли в условиях сильного течения.
Центр клетки занят вакуолью клеточного сока, цитоплазма занимает постенное положение. У мужоции (Mougeotia) (рис. 154: 1) хлоропласт в виде осевой пластинки с несколькими пиреноидами, у зигнемы (Zygnema) (рис. 154: 2) в цилиндрических клетках по два звездчатых осевых хлоропласта, каждый из которых содержит один крупный, центральный пиреноид.
280