

Кошкин
.pdfОценка относительной интенсивности потенциального фотосинтеза (определяли при концентрации 14СО2 1 %) различных органов сортов яровой пшеницы показала, что в сравнении с листовыми пластинками (интенсивность фотосинтеза которых принята за 100 %) интенсивность фотосинтеза влагалищ листа составляет от 39 до 47 %, открытой части междоузлия 30—39 % (Кумаков). Наибольшие различия между сортами наблюдаются в период от цветения до молочной спелости. Так, низкостебельные и полукарликовые сорта (Саратовская 52, Уорлд сидз 1616, Нададорес) отличаются повышенной ИФ колоса при существенно меньшем значении фотосинтеза соломины с листовыми влагалищами (табл. 2).
2. Вклад различных органов в общий фотосинтез верхнего участка побега сортов яровой пшеницы в период от цветения до молочной спелости, %
Часть побега |
|
|
|
|
|
|
выше стеблевого |
Альбидум43 |
Саратовская 29 |
Саратовская 52 |
Уорлд сидз 1616 |
Нададорес |
|
узла |
|
|
|
|
|
|
Листовая |
31 |
36 |
42 |
29 |
40 |
|
пластинка |
||||||
|
|
|
|
|
||
Влагалище листа |
26 |
20 |
17 |
18 |
21 |
|
Открытая часть |
21 |
21 |
11 |
9 |
2 |
|
междоузлия |
||||||
|
|
|
|
|
||
Закрытая |
|
|
|
|
|
|
влагалищем |
2 |
3 |
2 |
1 |
1 |
|
часть |
||||||
|
|
|
|
|
||
междоузлия |
|
|
|
|
|
|
Зерновки |
3 |
5 |
3 |
4 |
3 |
|
Остальные |
|
|
|
|
|
|
Части колоса |
16 |
15 |
25 |
39 |
33 |
|
(мякина) |
|
|
|
|
|
|
Колос в целом |
19 |
20 |
28 |
43 |
36 |
Определение интенсивности фотосинтеза целого растения пшеницы с использованием 14СО2 показало, что вклад стебля с влагалищем и колоса в общий фотосинтез после фазы трубкования был весьма значительным, достигая 70 % в фазе колошения и 90—95 % в фазе молочной спелости (в расчете на орган). Кроме того, фотосинтез колоса почти не подвержен полуденной депрессии, в то время как фотосинтез стебля и особенно листьев был сильно подавлен. В фазе молочной спелости у яровой пшеницы Саратовская 29 листья усваивают лишь около 17 %, колосья чуть больше 17 % и стебли с влагалищами листьев около 66 % всего СО2, поглощенного в процессе фотосинтеза целым растением.
Фотосинтезирующие органы существенно различаются и по степени использования их ассимилятов для формирования зерновок. Так, почти вся масса транспортных продуктов фотосинтеза
61

колоса попадает в зерновки, в то время как из верхнего листа — всего около 50 %, а из второго сверху листа — менее 20 %. Транспортные продукты фотосинтеза, притекающие из флага в колос, концентрируются в основном в зерновках, в то время как ассимиляты из листьев второго сверху яруса распределяются диффузно как в колосе, так и в зерновках, независимо от расположения колосков на колосковом стержне. В меченых (по С14) продуктах фотосинтеза стебля по сравнению с листьями доля радиоактивности доминирует в сахарозе при значительном уменьшении в органических фосфатах, кислотах и аминокислотах. Еще больший удельный вес сахарозы среди продуктов фотосинтеза колоса.
2.3.3. Чистая продуктивность фотосинтеза
Изучению чистой продуктивности фотосинтеза как важной слагающей урожая пшеницы уделено большое внимание. У этого показателя три основных достоинства: он определяется за длительные промежутки времени, в течение которых колебания, связанные с возрастным состоянием листьев, ярусной изменчивостью, кратковременными изменениями погодных условий и другими причинами, сглаживаются и элемент случайности сводится к минимуму; в изучение вовлекается большое число растений, что позволяет получать статистически достоверные данные; определения ЧПФ не требуют газометрических приборов, как при измерении интенсивности фотосинтеза, и более доступны в полевых условиях. ЧПФ характеризует собственно не фотосинтез в чистом виде, а суточную разницу между фотосинтезом и дыханием целого растения (не листа), отнесенную к единице листовой поверхности.
Расчет ЧПФ широко используют в исследованиях, так как он дает обобщенное и хорошо сопоставимое по вариантам, фазам развития, годам и т. д. представление об удельной производительности ассимиляционного аппарата
(рис. 20).
Рис. 20. Изменение ЧПФ по фазам развития яровой пшеницы Саратовская 29 за четыре разных года исследований
62
С улучшением питания и водоснабжения пшеницы повышаются интенсивность фотосинтеза и продуктивность растений, что сопровождается увеличением показателей ЧПФ. Вместе с тем между величиной ЧПФ и уровнем минерального питания не наблюдается четкой связи. Так, еще Watson (1947) отмечал положительное влияние полного минерального удобрения на площадь листьев пшеницы и — менее закономерное — на ЧПФ. Согласно этим данным, первый показатель имеет большую связь с урожаем, чем второй. Использование высоких доз удобрений и увеличение плотности посева, создаваемое загущением, как правило, снижают величину ЧПФ.
Этот негативный эффект можно свести к минимуму, улучшив оптикобиологическую структуру растений, в частности за счет изменения пространственного расположения листьев и уменьшения высоты стебля (см. с. 30). Низкорослые сорта с вертикальным расположением листьев характеризуются высокими величинами ЧПФ и Кхоз, так как такая структура благоприятствует хорошей освещенности ассимиляционной поверхности, оптимальной даже в период максимального развития листьев.
ЧПФ пшеницы не коррелирует с урожаями надземной массы и зерна. Более того, между величиной ЧПФ и урожаем биомассы наблюдается ясно выраженная обратная корреляционная зависимость (r= —0,76 ± 0,16). Это, видимо, связано с тем, что в фотосинтезе у пшеницы участвуют и другие ассимилирующие органы (стебель с влагалищами и колос), продукция которых приписывается листьям, что для такой культуры, как пшеница, сильно искажает результаты. В то же время массу корней, не говоря уже об опаде, корневых и летучих выделениях, как правило, не учитывают.
Рост урожайности селекционных сортов в засушливых условиях определяется главным образом увеличением площади листьев, ФП и Кхоз. При этом сохранение ЧПФ на одном уровне при увеличении размеров ассимиляционного аппарата в процессе селекции связано с малой площадью листьев. В связи с ограничением роста последней в этих условиях повышение ЧПФ следует рассматривать как главный резерв увеличения урожайности.
2.3.4.Пигментный комплекс и хлорофилльный индекс
Усортов озимой пшеницы наблюдается четкая дифференциация листовых пластинок по онтогенетической и ярусной изменчивости в содержании хлорофилла. Максимальное содержание хлорофилла отмечено в листовых пластинках в фазе колошение — цветение, минимальное — в фазе кущение (рис. 21).
63
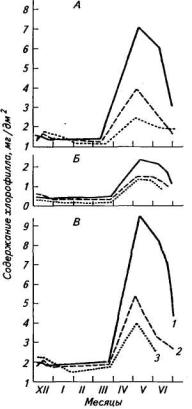
Рис. 21. Содержание хлорофиллов а (А), b (Б) и их сумма (В)
влистьях озимой пшеницы Безостая 1(1), Мироновская 808
(2)и Эритроспермум 7020 (3) в течение вегетационного
периода
Наибольшие различия в содержании хлорофилла между листовыми пластинками разных ярусов также наблюдались в фазе цветение, снижаясь в дальнейшем. Наиболее интенсивное разрушение хлорофилла было в листьях третьего сверху яруса, затем второго и флага.
Эффективность использования хлорофилла на фотосинтез у сортов озимой пшеницы зависит от ярусной и онтогенетической изменчивости листьев. Величины ассимиляционных чисел у листовых пластинок варьируют от 0,7 до 9,3 мг СО2/мг хлорофилла в час. Максимальные их значения наблюдаются в фазе трубкования во время активного
роста листьев. Верхние ярусы листьев отличаются большими величинами ассимиляционных чисел.
Эффективность использования хлорофилла на фотосинтез у влагалищ листа значительно ниже (0,5—3 мг СО2/(мг хл.∙ч), чем у
листовых пластинок и стебля (1,7—4), а в дальнейшем и колоса (2,3— 4,7). Что касается последнего, то эффективность использования хлорофилла в нем максимальна в фазе колошения, затем падает к молочно-восковой спелости.
Наиболее высокий коэффициент корреляции между содержанием пигментов в фотосинтезирующих органах и зерновой продуктивностью растений наблюдается в органах верхней зоны за вегетацию (r = +0,696), затем идут влагалища листьев (r= +0,626) и листовые пластинки (r = +0,612); причем в последних с урожаем зерна в большей степени коррелирует содержание хлорофилла а (r =+0,578), а с хлорофиллом b корреляционная зависимость слабая (r =+0,132). Существенная корреляционная связь наблюдается между урожаем зерна и содержанием в листовых пластинках каротиноидов (r= +0,660); содержание хлорофилла в фотосинтезирующих органах положительно коррелирует и с Кхоз.
64
Коэффициенты корреляции (r) между параметрами продуктивности и содержанием пигментов в различных органах пшеницы приведены ниже.
Коррелирующий |
Значение |
Коррелирующий |
Значение |
коэффициентов |
коэффициентов |
||
показатель |
корреляции (г) при |
показатель |
корреляции (г) |
ав |
Р005 - 0,468 |
бв |
при Р0>05 = 0,468 |
+0,696 |
+0,769 |
||
аг |
+0,626 |
бг |
+0,782 |
ад |
+0,612 |
бд |
+0,618 |
ае |
+0,578 |
бе |
+0,692 |
аж |
+0,132 |
бж |
+0,485 |
аз |
+0,660 |
|
|
Примечание, а — Ухоз, б — Кхоз, среднее за период содержание хлорофилла (а + b) в органах верхней зоны (в), во влагалищах листьев (г), в листовых пластинках (д), среднее за период вегетации содержание хлорофилла а (е), хлорофилла b (ж) и каротиноидов (з) в листовых пластинках озимой пшеницы.
Наиболее информативным признаком взаимосвязи содержания хлорофилла в отдельных органах и в целом растении с зерновой продуктивностью может быть не абсолютное его содержание, а соотношение с массой зерна. Этот показатель назван авторами коэффициентом хозяйственной эффективности хлорофилла.
Кхоз.эф.хл. = Ухоз/Хл,
где Ухоз — урожайность зерна, г/растение; Хл — содержание хлорофилла, мг/г сырой массы органа растения.
Данный показатель дает представление об эффективности использования хлорофилла на формирование зерновой продуктивности (в г/мг хролофилла). Коэффициент хозяйственной эффективности хлорофилла в большей степени коррелирует с зерновой продуктивностью, возрастая с ее увеличением. У высокопродуктивных сортов он значительно выше, несмотря на колебания в содержании пигмента (рис. 22). Это позволило рассчитать параметры продукционного процесса на единицу содержания хлорофилла в целом растении. Такой подход особенно оправдан по отношению к пшенице, у которой в фотосинтетической функции целого растения участвуют не только листовые пластинки, но и влагалища листьев, стебли и колосья. Обычно все расчеты привязывают к единице площади листьев, что неправильно.
В связи с этим для характеристики мощности развития фотосинтетического аппарата в агрофитоценозах пшеницы предложе-
65

Рис. 22. Коэффициент
хозяйственной эффективности |
Рис. 23. Структура ФП пшеницы |
|
Саратовская 29, рассчитанная по |
||
хлорофилла у различных сортов |
||
содержанию хлорофилла в |
||
озимой пшеницы: |
||
различных органах |
||
1 — Эритроспермум 7020; 2 — |
||
|
||
Кооператорка; 3 — Ульяновка; 4— |
|
|
Пиротрикс; 5—Лерма Рохо; б— |
|
|
Безостая 1; 7—Кавказ; 8— Гейнес; 9— |
|
|
Алма-Атинская 31; 10— Алма- |
|
|
Атинская полукарликовая; 11 — |
|
|
Днепровская 521 |
|
но определять не листовой индекс, а новую величину — хлорофилловый индекс, выражающийся в г хлорофилла/м2 или в кг хлорофилла/га агрофитоценоза. Для расчета этих параметров необходимо определять и суммировать количество хлорофилла во всех надземных органах растений. В связи с этим вместо понятия «фотосинтетический потенциал» предлагается использовать «хлорофилловый индекс» за весь вегетационный период или часть его (рис. 23).
Использование хлорофилловых фотосинтетических потенциалов (ХФП) по сравнению с поверхностными для оценки потенциальной продуктивности агрофитоценозов пшеницы имеет преимущество, поскольку в ХФП учитывают содержание пигментов не только в листьях, но и во всех надземных органах в онтогенезе. Выявлена тесная корреляционная связь хозяйственного урожая с величиной хлорофиллового фотосинтетического потенциала в благоприятных погодных условиях. Это свидетельствует о возможности использования данного признака в селекционной практике при оценке перспективных сортов пшеницы.
2.3.5. Коэффициент эффективности фотосинтеза
Поскольку продуктивность наряду с фотосинтезом определяется также и дыхательной компонентой, встает вопрос о взаимосвязи суммарного газообмена с накоплением биомассы у расте-
66
ний. Степень использования усвоенной СО2 на построение биомассы у растений характеризуется коэффициентом эффективности фотосинтеза (K^).
Он показывает, какое количество сухой биомассы растение образует в течение суток при усвоении единицы (1 г, 1 кг) СО2. Теоретически возможный К^ равен 0,68. В благоприятных условиях значение Кэф может приближаться к 0,5, а в неблагоприятных — снижаться до нуля или даже быть отрицательным. Чаще всего величины Кэф колеблются, значительно изменяясь в онтогенезе. Так, у яровой пшеницы Кэф был максимальным в фазе кущения — 0,42, а в фазе молочной спелости он снизился до 0,08. Следовательно, эффективность фотосинтеза от колошения до начала молочной спелости снизилась более чем в 5 раз. У озимой пшеницы Кэф в среднем за вегетацию составил 0,4-0,42.
Изменчивость Кэф в ходе вегетации может зависеть также от неполного учета биомассы в результате потерь веществ, выделенных растениями через корни в процессе экзоосмоса, от образования гелеобразных корнеоблегающих чехлов и от летучих газообразных выделений. По расчетам С. А. Самцевича, растения озимой пшеницы и кукурузы за счет образования гелеобразных веществ корнеоблегающих чехлов могут выделить в почву 70—125 ц/га сухого вещества. Величина потерь определяется длиной вегетационного периода и условиями выращивания.
В итоге это приводит к значительным колебаниям ЧПФ и определяет несоответствие между интенсивностью фотосинтеза, его суточной продуктивностью и биологическим урожаем, а также между последними и ЧПФ.
2.3.6. Коэффициент использования солнечной энергии (КПД ФАР)
КПД приходящей ФАР на формирование урожаев яровой и озимой пшеницы в различных зонах страны в среднем за период вегетации варьирует от 0,3 до 4 %(табл. 3). В острозасушливые годы КПД ФАР падает до 0,3 %; при достаточной влагообеспеченности на фоне различного минерального питания он составляет 1—3 %, а при сочетании орошения с минеральным питанием — 3—4 %. С ростом КПД ФАР, как правило, увеличиваются биологические и хозяйственные урожаи. Однако при высоких урожаях биомассы Кхоз несколько снижается. При этом наблюдается обратная зависимость между величинами КПД ФАР и Кхоз.
67
3. Использование солнечной энергии (ФАР) посевами пшеницы и ее продуктивность
КПД ФАР |
Убиол, ц/га |
Ухоз, ц/га |
Кхоз |
Автор |
|
|
Яровая пшеница |
|
|
0,30 |
17,6 |
4,0 |
0,24 |
Кумаков, 1971 |
1,50 |
89,8 |
35,7 |
0,43 |
То же |
3,30 |
132,0 |
39,9 |
0,30 |
Устенко, 1969 |
4,07 |
186,0 |
57,4 |
0,31 |
Тоже |
|
|
Озимая |
пшеница |
|
0,90 |
58,8 |
18,5 |
0,31 |
Клименко и др., 1973 |
1,50 |
98,5 |
32,4 |
0,39 |
То же |
2,83 |
141,0 |
49,3 |
0,35 |
Островская и др., 1969 |
3,43-3,67 |
215,0-230, |
0 67,3-69, |
4 0,25-0,28 |
Ивакин, 1970 |
Примечание. Убиол — урожай биологический, Ухоз — урожай хозяйственный.
У высокопродуктивных сортов интенсивного типа селекции академика П. П. Лукьяненко, имеющих высокий выход зерна (0,4— 0,53) и сохраняющих его при возрастающих дозах удобрений и водообеспеченности, максимальные урожаи зерна в 70—80 ц/га и даже более 90 ц/га наблюдаются обычно при биологических урожаях 160—180 ц/га, т. е. при КПД ФАР 3—4 %. Такие сорта обладают большим потенциалом урожайности и при дальнейшем увеличении КПД ФАР и общей биомассы могут дать значительную прибавку.
Теоретически можно получать урожаи с КПД приходящей ФАР 7—8 % и даже 10 % (при оптимальном снабжении водой и элементами минерального питания). Это значит, что в районах, где приход ФАР за период вегетации составляет 2—3 млрд ккал/га, урожай зерна пшеницы может достигать 150 ц/га. О реальности таких урожаев свидетельствуют примеры создания в отечественной и мировой селекции низкорослых (короткостебельных) сортов пшеницы интенсивного типа, дающих 100 ц/га и выше. Что касается теоретически возможных КПД ФАР, то они составляют 28—29 % по газообмену и 20—22 % по запасаемой в биомассе энергии.
Таким образом, фактически наблюдаемые коэффициенты использования энергии в посевах на фотосинтез и формирование урожаев в подавляющем большинстве случаев далеки от оптимальных величин и тем более от теоретически возможных. Это свидетельствует о значительных резервах увеличения урожайности растений.
2.3.7. Фотосинтез и урожай пшеницы
Представление о связи фотосинтеза растений с урожайностью первоначально базировалось на том, что между ними существует прямая зависимость. Поэтому проводились исследования по установлению положительной связи между интенсивностью фотосин-
68

теза и общей фитомассой, в том числе хозяйственным урожаем (Тимирязев, 1937).
Однако в дальнейшем было установлено, что эта зависимость гораздо сложнее. Наиболее глубоко этот вопрос был проработан Л. А. Ивановым (1941), выразившим зависимость между урожаем, фотосинтезом и дыханием простым уравнением, которое стало классическим:
М + т =fРТ - аР'Т',
где М — общая сухая фитомасса всего растения (без азота и золы); т — масса опадав— интенсивность фотосинтеза; Р — фотосинтезирующая площадь; Т —продолжительность процесса фотосинтеза; а — интенсивность дыхания; Р' —дышащая масса растений; Т' — продолжительность процесса дыхания; fРТ — продуктивность фотосинтеза. Интенсивность фотосинтеза и дыхания выражена в эквивалентах биомассы.
Согласно этому уравнению, урожай зависит в первую очередь от продуктивности фотосинтеза, а продуктивность — от интенсивности фотосинтеза, размера рабочей поверхности и рабочего времени за период учета урожая.
В дальнейшем А. А. Ничипорович внес существенные дополнения в формулу Л. А. Иванова. Он предложил уравнение связи между фотосинтезом и накоплением органического вещества в динамике формирования не только биологического (Убиол), но и хозяйственного (Ухоз) урожаев в агрофитоценозе. А. А. Ничипорович в формулу ввел новые важные элементы уравнения — коэффициент эффективности фотосинтеза (Кэф) и коэффициент хозяйственной эффективности (Кхоз):
где ФСО2 — дневная продуктивность фотосинтеза; Л — площадь или индекс листовой поверхности (ИЛП); Кэф — коэффициент эффективности фотосинтеза; Кхоз — коэффициент хозяйственной эффективности; п — время активной работы фотосинтетического аппарата.
Таким образом, наивысшие урожаи могут быть получены при интенсивном росте размеров площади листьев растений в агрофитоценозе, при наибольшем времени (n) активной работы фотосинтетического аппарата в течение каждых суток и вегетационного периода, т. е. при наиболее высоких значениях фотосинтетических потенциалов (ФП), при наиболее высоких суммах дневного усвоения СО2, а также высоких коэффициентах эффективности фотосинтеза (Кэф).
Итак, все агротехнические мероприятия, включая применение удобрений, поливы и т. д., а также селекционно-генетические ме-
69
роприятия должны быть направлены на поддержание оптимального хода указанных процессов и согласования процессов фотосинтетического и дыхательного метаболизма.
2.3.8. Продукционный процесс зерновых культур при повышенной концентрации СО2 в воздухе
Предполагается, что содержание в атмосфере СО2 0,035 %, которое наблюдалось в конце XX в., ко второй половине XXI в. удвоится и достигнет 0,07 %. Подобное изменение климата вызовет изменение донорно-акцепторных отношений и размеров ассимиляционной поверхности, повысит потенциал продуктивности растений, изменит потребность сельскохозяйственных культур в минеральном питании.
Фотосинтез и рост пшеницы при повышенной концентрации СО2 в
воздухе. Степень увеличения фотосинтеза в ответ на повышение концентрации СО2 в атмосфере зависит от внешних (температура, водообеспеченность, освещенность, минеральное питание) и внутренних (устьичная проводимость, карбоксилирующая активность, напряженность донорно-акцепторных отношений в растении и др.) факторов.
При длительном произрастании пшеницы при повышенной концентрации СО2 эффект от начальной активации фотосинтеза (29—98 %) и ингибирования дыхания (40—60 %) исчезает, и такие растения начинают отставать в росте от растений, произрастающих при нормальной концентрации СО2. Одна из причин этого — уменьшение эффективности работы (или скорости регенерации) рибулезобисфосфат-карбоксилазы/оксигеназы (РБФК/О). Теоретически интенсивность карбоксилирующей работы РБФК/О может повышаться при увеличении СО2 на 20—30 % при одновременном торможении оксигеназной активности на 40 %. Однако на практике при длительном воздействии высокой концентрации СО2 на посевы пшеницы наблюдается уменьшение активности РБФК или ее количества. Причиной этого может быть снижение интенсивности синтеза самого белка РБФК/О в результате ограничения доступности азота и недостатка АТФ.
В первой половине вегетации (10—13 дней) в атмосфере с повышенным содержанием СО2 наблюдается активизация фотосинтеза пшеницы (на 29— 98 %) при одновременном снижении дыхания (на 42—56 %). Результатом этого являются увеличение величины нетто-фотосинтеза и соответственно активизация роста при одновременном увеличении площади листьев растения. Высокий уровень минерального, в частности азотного, питания позволяет при этом сформировать в 2 раза большую площадь листьев.
Реакция растений на повышение СО2 в более поздний период вегетации слабее. Высокие температуры, активизируя дыхание растений, способствуют увеличению соотношения дыхания и фото-
70