

Кошкин
.pdfпам органогенеза можно со сравнительно высокой степенью достоверности судить о том, в каком возрастном периоде и в какой фазе развития находится растение.
Вразвитии однолетнего побега пшеницы выявлено 12 этапов органогенеза. Рассмотрим основные из них.
Первый этап формирования побега пшеницы начинается с образования инициальных клеток промеристемы. Из этих клеток формируется первичная образовательная ткань конуса нарастания — меристема, которая дает начало зачаткам зародышевых листьев и зародышевого стебля. Этот этап наблюдается
впериод фенофазы всходов и определяет следующие элементы продуктивности: прорастание, всхожесть, густоту стояния растений.
Второй этап связан с фенофазой третьего листа и кущением. Он характеризуется дифференциацией основания конуса нарастания на зачаточные узлы, междоузлия и зачатки стеблевых листьев. Это определяет габитус растения (высота, число листьев), коэффициент кущения, зимостойкость. В этой фенофазе наблюдается и третий этап органогенеза — процесс дифференциации главной оси зачаточного колоса и зачаточных кроющих листьев и образование числа члеников колоса.
Вфенофазе начала выхода в трубку наблюдается четвертый этап — образование конусов нарастания второго порядка (колосковых бугорков), что определяет число колосков в колосе и продуктивную кустистость.
Вфенофазе выхода в трубку — начала стеблевания происходят: пятый этап органогенеза — закладка покровных органов цветка, тычинок и пестика, определяющих число цветков в колосках, шестой этап — формирование соцветия и цветка (микро- и макроспорогенез) и седьмой этап — гаметофитогенез, рост покровных органов, удлинение члеников колоскового стержня, определяющих фертильность цветков и плотность колоса.
Вфенофазе колошения наблюдается восьмой этап органогенеза — гаметогенез, завершение процессов формирования всех органов соцветия и цветка.
Вфенофазе цветения проходят девятый этап — оплодотворение и образование зиготы и десятый этап — рост и формирование зерновки, определяющие озерненность колоса и величину зерновки.
Вфенофазах молочной и восковой спелости наблюдаются одиннадцатый этап — накопление питательных веществ в зерновке (семени) и двенадцатый этап — превращение питательных веществ в запасные вещества в зерновке (семени), определяющие массу зерновки.
Таким образом, в ходе онтогенеза у растений пшеницы одновременно протекают возрастные, этапные и органообразующие процессы. В то же время анализ взаимосвязей этих процессов показывает, что нормальный морфогенез органов — одно из важнейших внутренних условий для осуществления возрастных и
51
стадийных процессов в онтогенезе растения. Без образования соответствующих морфоструктур с характерными для них физиологическими функциями и качественно новыми превращениями веществ нормальный ход онтогенетических процессов задерживается и, несмотря на наличие благоприятных условий, зачастую прекращается.
2.1.2. Строение, рост и ярусная изменчивость листьев
Листья пшеницы разделяют на зародышевые, прикорневые и стеблевые. При формировании зародыша в семени из поверхностного слоя меристемы конуса нарастания образуются листовые бугорки. Разрастание первого листового бугорка приводит к образованию первого листа. Затем таким же способом в определенной последовательности образуются второй и третий зародышевые, или прикорневые, листья.
Рост листьев и развертывание листовых пластинок происходят в период от появления всходов и до IX этапа органогенеза.
Всего на главном побеге у большинства сортов пшеницы закладывается 8—10 листьев; на боковых побегах кущения в зависимости от порядка кущения образуется на 1—3 листа меньше. При интенсивном кущении на одном растении может образоваться 30—40 побегов, несущих по 3—6 листьев, общее число листьев превышает 100—120.
Листья различных ярусов растений пшеницы различаются по многим анатомическим признакам. Наиболее четко различия проявляются при учете величины и количества устьиц. Нижние листья имеют меньше устьиц по сравнению с верхними. При этом на верхней стороне листовых пластинок их всегда несколько больше, чем на нижней. Влагалища нижних листьев имеют значительно меньше устьиц по сравнению с листовыми пластинками, влагалища верхних листьев с наружной стороны образуют почти столько же устьиц, как и их листовые пластинки.
Во влажные годы устьиц меньше, чем в сухие. Листья разных ярусов различаются как по числу устьиц, так и по числу и величине сосудистых пучков. У нижних листьев (второго и третьего снизу ярусов) число крупных сосудистых пучков колеблется в зависимости от сорта от 16 до 24, а у верхних листьев — от 20 до 32.
На II этапе онтогенеза все развернувшиеся листья очень сходны как по строению, так и по выполняемым функциям. Обычно у них очень слабо развито листовое влагалище, поэтому эти зародышевые листья называют прикорневыми.
Над влагалищем листа (у основания листовой пластинки) расположен вырост, или язычок, имеющий два выступа, которые называют ушками. Лишь у небольшой группы безлигульных пшениц ушки не развиваются.
52
Изучение анатомического строения тканей листьев пшеницы показало, что все сосудистые пучки имеют коллатеральное строение: ксилема направлена к верхней поверхности пластинки, флоэма — к нижней. Большие пучки с сильно развитой ксилемой выполняют преимущественно функции водоснабжения. Малые пучки, состоящие главным образом из флоэмы, участвуют в сборе и оттоке продуктов ассимиляции. Малые пучки сосредоточены в основном в верхних листьях пшеницы.
Лист пшеницы способен изменять направление своего расположения к лучам солнца, так что в большинстве случаев верхняя сторона той или иной его части поглощает не только отраженный, но и прямой солнечный свет. При этом листья верхних ярусов за 1 сут описывают почти полный круг. Такое изменение ориентации листовой пластинки позволяет листьям в засушливых условиях защищаться от перегрева, а во влажных районах лучше использовать свет и тепло, которые там находятся в минимуме.
В связи с этим большой интерес представляет гибрид озимой пшеницы Гостианум 88, обладающий свойством верхнего (флагового) листа после выхода колоса опускаться и прижиматься к стеблю, что способствует осветлению агрофитоценоза, уходу листа от интенсивной инсоляции в жаркие часы дня. Это положительно сказывается на продуктивности растений.
Наибольшее изменение в направлении расположения листьев у пшеницы наблюдается при их вертикальной ориентации в сочетании с низкоросл остью растений.
Рост листьев разных ярусов коррелятивно связан с ростом одноименных междоузлий и прохождением растением определенных этапов органогенеза. При нормальных условиях выращивания по числу листьев, вышедших из влагалища предыдущего листа, можно со сравнительно большой степенью приближенности судить об этапе органогенеза, на котором в тот или иной момент находится растение.
Такое проявление связи между развитием побега, состоянием его генеративных и ростом вегетативных органов (листьев и междоузлий стебля) обусловлено связью между ярусом листа, его физиологическими функциями и той ролью, которую играет каждый лист в процессах онтогенеза и формирования репродуктивных органов.
2.1.3. Строение, рост и ярусная изменчивость междоузлий стебля
Стебель, или соломина, злаков — основной осевой орган, он несет листья и колос. Стебель выполняет механическую (опорную) и проводящие функции, состоит из метамерных (последовательно расположенных) органов — узлов с листьями и междоузлий,
53
Метамеры стебля формируются благодаря деятельности верхушечной меристемы на I и II этапах органогенеза. Дифференциация стебля на метамеры начинается еще в зародыше, когда формируются зародышевые листья. Уже на II этапе органогенеза (до начала видимого роста стебля) можно видеть узлы и междоузлия стебля. Однако в сельскохозяйственной практике начало роста стебля принято определять с момента удлинения первого надземного междоузлия, т. е. с фазы выхода в трубку.
Рост стебля у пшеницы выражается главным образом в значительном удлинении междоузлий и в меньшей мере в их утолщении. Интеркалярный рост междоузлий в длину осуществляется в результате деления меристематических клеток и последующего их растяжения.
Для роста стебля, как и растения в целом, необходимы определенные условия освещения, длина дня, температура и другие факторы, обеспечивающие развитие пшеницы.
При малейшем дефиците влаги или резком понижении температуры рост очередного междоузлия приостанавливается и оно остается укороченным. Наиболее интенсивно междоузлия стебля растут при температуре 24—25 оС.
2.1.4. Строение, рост и развитие корневой системы
Корневая система пшеницы, как и других однолетних злаков, мочковатая. В период прорастания и появления всходов образуется первичная корневая система, а затем в начале процесса кущения развивается и вторичная (рис. 17).
Как первичные, так и вторичные корни пшеницы образуют корневые волоски, которые размещаются на расстоянии 0,1—1,2 см от кончиков корешков. Та часть корневой системы, которая покрыта корневыми волосками, называется активной деятельной областью. Общая поверхность корневых волосков у одного пшеничного растения очень велика (до нескольких квадратных километров), благодаря чему корневая система растений может использовать минимальные количества минеральных элементов, находящихся в почве.
Волоски корешков имеют вид закрытых трубок длиной 1,5— 2 мм. Почвенный раствор, проникая в полость волосков, пропитывает стенки, образует слизевой слой, благодаря которому частицы почвы легко пристают к корешкам.
Рост корней в длину опережает рост надземных органов. У яровой пшеницы уже через 10—15 дней после всходов длина корней превышает 45— 50 см, а еще через 15—20 дней достигает 80—90 см. У озимой пшеницы при посеве осенью корни перед уходом растений в зиму достигают глубины 100 см.
Корневая система, определяющая водный режим и минеральное питание растений, наиболее тесно связана с формирванием
54

Рис. 17. Образование нескольких ярусов корней из узлов главного побега у пшеницы (цифрами указан порядок ярусов боковых корней, бп — боковые побеги)
зерновой продуктивности пшеницы. Известно, что урожай яровой пшеницы можно получить и тогда, когда растения развивают лишь зародышевые корни. Они играют особенно большую роль в засушливые годы, когда из-за значительного дефицита влаги в поверхностных слоях почвы узловые корни недоразвиваются и растения растут почти исключительно за счет зародышевых корней; их деятельность в основном и ограничивает величину урожая.
При оптимальном водоснабжении растений и особенно в условиях периодического орошения значительно повышается вклад узловых корней в получении высокого урожая, который зависит от наличия не только зародышевых, но и в большей степени узловых корней. При типичной структуре корневой системы в засушливых степных районах (состоящей преимущественно из зародышевых корней) на долю первых трех зародышевых корней в создания урожая приходится 30—40 %, на долю второй пары за-
55
родышевых корней — 20—30, колеоптильных — 20—30 и на долю узловых корней лишь от 5 до 30 %. Совершенно иное положение складывается при достаточном увлажнении в условиях высокого фона плодородия почвы, когда урожай достигает 50—60 ц/га. Узловые корни, глубоко проникая в почву, участвуют в создании более 40—50 % от общего урожая. В этом случае основное значение имеет раннее развитие первичной корневой системы.
Имеются данные, что у пшеницы и ячменя наблюдается тесная прямая связь (коэффициент корреляции близок к единице) между урожаем надземной массы растения и числом узловых корней. В связи с этим нашел практическое применение метод отбора селекционного материала по мощности корневой системы на повышение урожая и улучшение его качества у озимой пшеницы.
2.2. Строение зерновки и физиология прорастания семян
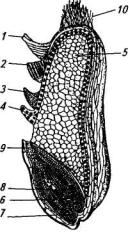
Строение зерновки. Зерновка пшеницы — сухой односемянный нераскрывающийся плод злаков, который в агрономической практике обычно называют зерном. В зерновке следует различать собственно семя, состоящее из зародыша, эндосперма и семенных оболочек, и плодовую оболочку, представляющую собой стенки завязи (рис. 18). Размеры зерновки в зависимости от вида, сорта и условий произрастания сильно изменяются. Длина ее колеблется от 4 до 9 мм, ширина —от 0,8 до 2 мм и толщина —от 1,5 до 3,5 мм. Различаются они также и по массе — от 20 до 90 мг.
Зародыш расположен у основания зерновки под некоторым углом к эндосперму и может быть легко отделен от него. Он состоит из щитка, который соединяет его с эндоспермом, почечки и зачаточных бугорков корешков. Зародышевая почка семени состоит из конуса нарастания, первичного зачаточного стебля и зародышевых листьев, закрывающих в виде колпачка конус нарастания.
Средняя масса зародыша колеблется от 1,7 до 3% общей массы зерновки, масса плодовых и семенных оболочек — от 11,3 до 17,3 и масса эндосперма — от 78,5 до 87 %.
В некоторых случаях в одной зерновке образуются два или даже три зародыша. Это
Рис. 18. Продольный разрез зерновки пшеницы:
1—3— плодовые и семенные оболочки; 4—алейроновый слой; 5— эндосперм; б—зародыш 7—зачаточный корешок; 8—почечка; 9— щиток; 10— бородка
56
явление чаще всего наблюдается при выращивании пшеницы на высоком агротехническом фоне и при избытке азота на V—IX этапах органогенеза. Явление беззародышевости зерновок, когда при нормально развитом эндосперме зародыш отсутствует, встречается редко. Еще реже наблюдается нормальное развитие зародыша при редуцированном эндосперме.
В эндосперме можно выделить наружный (алейроновый) слой, который почти не содержит крахмала, и собственно эндосперм, клетки которого содержат крахмальные зерна.
Прорастание семян. Оптимальные условия для прорастания зерновок пшеницы наступают, когда они поглощают 45—47 % воды (в % к воздушносухой массе). Набухание зерновок пшеницы сначала идет за счет поглощения воды, а к концу его действуют осмотические силы, возникающие в результате гидролиза полисахаридов и других веществ. При этом ксерофитные сорта нуждаются при прорастании в меньших количествах воды, чем мезофитные.
Скорость прорастания, появление корешка, а затем и дальнейший рост зародыша зависят при достаточном насыщении водой также от температуры. Уже при температуре таяния льда зерновки пшеницы могут поглощать воду. Так, по данным А. И. Носатовского, зерновки пшеницы в песке при влажности 90 % и температуре таяния льда за 15 ч поглощали до 30 % влаги от массы сухих зерновок. С повышением температуры при влажности песка 90 % скорость поглощения воды резко возрастает. Так, при температуре 24 °С через 1 сут зерновки содержали 50 % влаги. Оптимальные температуры для прорастания семян пшеницы колеблются в зависимости от сорта и влажности семян от 18 до 25 °С, минимальные — от 1 до 2 °С.
На скорость прорастания семян влияет доступ кислорода. Затрудненный доступ кислорода при увеличении глубины посева семян или избыточном содержании воды в почве снижает их всхожесть. С началом роста зародыша потребление кислорода резко увеличивается. Диоксид углерода (СО2), напротив, снижает энергию прорастания семян.
Любое повреждение зерновки отрицательно влияет на всхожесть и дальнейший рост растений. Так, удаление в зерновках призародышевой части эндосперма со спинки вызывало у проростков не только потерю геотропической ориентации, но и глубокие физиологические изменения: резкое падение интенсивности фотосинтеза, ослабленное развитие корневой системы, значительное отставание процессов дифференциации зачаточного конуса нарастания (Куперман).
Нормальное поглощение воды обеспечивает деятельность многочисленных ферментов в набухающих и прорастающих зерновках. Под влиянием амилазы (состоящей из α- и β-амилазы) крахмал в эндосперме зерновки переводится в декстрины и мальтозу.
57
Мальтоза под влиянием мальтазы расщепляется при прорастании семян до глюкозы. Одновременно с накоплением глюкозы идет образование сахарозы, которая используется растущим проростком.
Изучение протеолитических ферментов зерна показало, что резкое повышение активности протеиназы наблюдается также при прорастании зерна. При расщеплении белковых веществ количество амидного азота увеличивается, но общее содержание его при прорастании остается на одном уровне. Лишь спустя 12—15 дней количество азота в проростках начинает увеличиваться за счет усвоения его из почвенного раствора.
Таким образом, при наличии необходимых условий (соответствующей влажности ложа, температуры и доступа кислорода) семена пшеницы, завершившие XII этап органогенеза, способны нормально прорастать и вновь начинать жизненный цикл с I этапа органогенеза.
2.3. Фотосинтетическая деятельность и фотосинтетическая продуктивность пшеницы
2.3.1. Фотосинтетический потенциал
Ассимиляционная поверхность пшеницы определяется не только листовыми пластинками, но и другими ассимилирующими хлорофиллоносными органами (соломина с листовыми влагалищами и колос). В. А. Кумаковым был предложен метод оценки вклада полиассимилирующих органов в фотосинтетической деятельности растений пшеницы. Он основан на анализе структуры их фотосинтетического потенциала (ФП), т. е. суммарной ассимиляционной поверхности за отдельные периоды и за всю вегетацию. Основанием для применения этого метода служит тот факт, что из всех фотосинтетических показателей урожай растения теснее всего коррелирует с мощностью ассимиляционного аппарата, т. е. с величиной фотосинтетического потенциала.
Если результаты определения ФП отдельных органов изобразить графически (рис. 19), то можно ясно представить состав работающего ассимиляционного аппарата за любой отрезок времени. Для этого необходимо опустить перпендикуляры к оси абсцисс из точек, соответствующих началу и концу интересующего нас периода, и определить площади фигур, отображающих фотосинтетические потенциалы разных органов и заключенных между перпендикулярами.
В таблице 1 показана доля определенных листьев в общем ФП всех листьев за вегетацию у сорта яровой пшеницы Саратовская 29. Если отбросить девятый лист, появляющийся очень редко, то почти 60 % всего ФП листьев за вегетацию приходится на три верхних листа, а за период после колошения 2/3 всей работы листьев выполняют два верхних листа.
58

Рис. 19. Структура ФП яровой пшеницы сорта Альбидум 43: 1—8—листья; 9— соломина; 10— колос
1. Вклад отдельных органов растений яровой пшеницы Саратовская 29 в фотосинтетический потенциал главного побега в различные периоды вегетации
|
|
ФП отельных органов (см2∙сут) по периодам |
|||||
Ассимилирующий |
всходы |
кущение |
выход в |
колошение |
начало |
всего за |
|
— начало |
молочной |
||||||
орган |
— |
— выход |
трубку — |
вегетационный |
|||
|
кущение |
в трубку |
колошение |
молочной |
спелости |
период |
|
|
спелости |
— уборка |
|||||
Лист: 1-й |
48 |
37 |
11 |
— |
— |
96 |
|
2-й |
42 |
37 |
21 |
— |
— |
100 |
|
3-й |
17 |
66 |
69 |
— |
— |
152 |
|
4-й |
6 |
74 |
116 |
3 |
— |
199 |
|
5-й |
— |
43 |
204 |
51 |
6 |
304 |
|
6-й |
— |
8 |
205 |
113 |
25 |
351 |
|
7-й |
— |
— |
177 |
138 |
115 |
430 |
|
8-й |
— |
— |
99 |
111 |
121 |
331 |
|
9-й |
— |
— |
7 |
10 |
10 |
27 |
|
Все листья |
113 |
265 |
909 |
426 |
277 |
1990 |
|
Соломина |
— |
— |
301 |
342 |
770 |
1413 |
|
Колос |
— |
— |
— |
361 |
669 |
1030 |
|
Растение в целом |
113 |
265 |
1210 |
1129 |
1716 |
4433 |
|
При оценке роли разных органов, с учетом соломины и колоса, установлено, что в целом за вегетацию на долю листовых пластинок приходится менее половины всего ФП главного побега, а за период от колошения до уборки доля листьев лишь немного превышает 1/4 ФП побега. При этом участие листьев в ФП побега мало изменяется по годам, тогда как относительный вклад соломины и
59
колоса варьирует очень сильно: в сухие годы повышается значение колоса, а во влажные — соломины с листовыми влагалищами. Изучение вклада отдельных листьев, соломины и колоса в ФП главного побега в отдельные периоды вегетации у яровой пшеницы Саратовская 29 показало, что до кущения главную роль играют два нижних листа, от кущения до выхода в трубку — 3—5-й листья, от выхода в трубку до колошения потенциал листьев максимален при преимущественной роли 4—7-го листьев, от колошения до молочной спелости листья (в основном 6—8-й) играют еще более заметную роль, а в заключительный период вегетации работают лишь два листа, основная роль отводится соломине и колосу.
2.3.2. Интенсивность фотосинтеза листьев и других хлорофиллоносных органов
Изучению интенсивности фотосинтеза листьев пшеницы как главного фактора формирования урожая посвящено много исследований. Основное внимание в них уделено дневным и сезонным изменениям фотосинтеза в связи с формированием биологического и хозяйственного урожаев, а также влиянию условий возделывания и сортовых особенностей на интенсивность процесса.
Изучая интенсивность фотосинтеза пшеницы в ходе вегетации в контролируемых условиях, А. М. Osman и F. L. Milthorpe установили, что интенсивность фотосинтеза и дыхания увеличивалась с возрастом растения к фазе выколашивания, после чего снижалась (интенсивность дыхания медленнее, чем фотосинтеза). В. А. Кошкин и О. Д. Быков наблюдали два максимума интенсивности фотосинтеза: первый — конец фазы выхода в трубку
—начало колошения, второй — фаза цветения — налив зерна.
Вполевых неконтролируемых условиях интенсивность фотосинтеза постепенно нарастает от начала развития растений до их перехода к репродукции (колошению), где достигает своего максимума, а затем начинает снижаться. По мнению Ф. Д. Сказкина, наибольшая интенсивность фотосинтеза наблюдается в период формирования микроспор. Затем в период цветение — начало формирования зерна интенсивность фотосинтеза, как правило, несколько снижается, а потом сменяется подъемом во время налива зерна и спадом в конце вегетации, наиболее резким у скороспелых сортов и более плавным у среднеспелых. Интенсивность фотосинтеза у позднеспелых сортов сохраняется более продолжительное время.
Максимум ИФ у колоса отмечен в период формирования и налива зерна, и его величина в расчете на 1 г составляла 36,6 мг СО2, или 9,7 мг на один колос. В этот период у колоса, масса которого была 267 мг, величина ассимиляции СО2 за один день равнялась примерно 100 мг.
60