

Кошкин
.pdfиной мере может ограничивать урожай. Например, урожай пшеницы не реагирует на повышенную концентрацию СО2 при пониженных температурах воздуха ни в какой фазе развития, но возрастает при высоких температурах. Улучшение освещенности верхней трети профиля посева высокоурожайных гибридов кукурузы с помощью специальных отражателей перед цветением початка и сразу после него увеличивает урожай сильнее, чем во время роста зерновок, главным образом благодаря увеличению числа зерен в початке и массы апикальных зерен. Подобные результаты получены также при прореживании посевов кукурузы и пшеницы в поле, что улучшает радиационный режим и соответственно продуктивность одного растения.
1.1.3. Лимитирование акцептором
Уменьшенный акцептор. Снижение размеров акцептора ниже определенного уровня, как правило, снижает урожайность. Однако одновременно может происходить некоторое компенсаторное увеличение элементов урожайности, детерминируемое на более поздних фазах развития. Поэтому интерпретация случаев с удалением части акцептора также осложняется контролем фотосинтеза по принципу обратной связи, приводящим к снижению ИФ или ускоренному старению листьев.
Нормальный баланс. Одним из доказательств того, что донор не полностью лимитирует урожай, является наличие зачастую достаточно значительного количества резервов углеводов в растениях при уборке, как, например, у кукурузы, выращиваемой на низких широтах. Другой не менее интересный факт — часть листьев все еще остается зеленой и фотосинтетически активной, когда рост зерновки уже тормозится, как, например, у пшеницы и кукурузы. Ингибирование фотосинтеза по принципу обратной связи, как правило, предотвращает подобную ситуацию. Однако наличие указанного факта предполагает, что продолжительность функционирования акцептора у некоторых культур по меньшей мере лимитирует урожайность. Кроме того, тот факт, что ИФ культуры часто увеличивается с усилением роста генеративных органов, предполагает, что при наличии большей аттрагирующей способности донор мог бы быть более активным.
Усиленная активность акцептора. Как отмечалось ранее, повышение концентрации СО2 или освещенности увеличивает урожай сильнее всего при их действии до фазы цветения. Именно тогда определяется потенциальная запасающая емкость культуры, зависящая также от генотипа. Влияние генотипа лучше всего можно продемонстрировать в опытах с прививками, особенно клубне- и корнеплодов. Например, размер клубня картофеля и содержание крахмала не зависели от массы ботвы, т. е. урожай был лимитиро-
11
ван преимущественно активностью акцептора. Этот вывод подкреплен также при прививке низкоурожайного сорта сладкого картофеля на высокоурожайный. Эти опыты показали широкий спектр активностей донора и акцептора и их соотношений. При этом у одних сортов обнаружена высокая активность донора и низкая — акцептора, а у других — наоборот. Урожай теснее коррелирует с активностью акцептора, чем донора. Однако для получения высоких урожаев необходима высокая активность как донора, так и акцептора.
Приведенные данные показывают, что как донор, так и акцептор не являются независимыми детерминантами урожая и оба могут в определенной мере его лимитировать. Некоторые модели продукционного процесса допускают это. При этом предполагается, что единый общий пул сахаров контролирует как ИФ листа, так и скорость роста плодоэлемента. Если масса последнего растет с повышением концентрации сахаров в пуле на фоне снижения ИФ, данная культура не может функционировать при полной активности донора и акцептора, так как уровень концентрации сахаров, увеличивающий одну скорость, минимизирует другую. Однако уровень сахаров в листьях может и не быть фактором, регулирующим фотосинтез, и поскольку загрузка флоэмных окончаний является активным процессом, низкое содержание сахаров в фотосинтезирующих тканях может быть связано со значительно более высоким их уровнем в запасающих тканях. Поэтому предположение о существовании общего пула сахаров для донора и акцептора представляется маловероятным.
Таким образом, причины того, что донор и акцептор одновременно лимитируют урожай, можно искать везде. Лимитирование донором и акцептором может меняться даже в течение суток по мере изменения освещенности, влияющей на ИФ (донор), и температуры, воздействующей на рост плодоэлемента (акцептор). У современных районированных сортов, адаптированных к условиям выращивания, донорно-акцепторные отношения, видимо, достаточно хорошо сбалансированы, за исключением случаев действия экстремальных биотических и абиотических факторов. Поэтому дальнейшее увеличение потенциала урожая возможно за счет одновременного усиления активности как донора, так и акцептора. Рассмотрим сначала изменение активности донора.
1.2. Фотосинтез
Повышение ИФ возможно как генетическим путем, так и благодаря совершенствованию агротехники, хотя этого может быть и недостаточно для увеличения урожайности. Существует заблуждение, что фотосинтез неэффективен, так как только малая доля падающей радиации используется на формирование урожая, а также
12
потому что есть некоторые процессы, в частности фотодыхание, которые просто бесполезны. На самом деле при своей максимальной эффективности поглощения солнечной энергии процесс фотосинтеза сравним с лучшими фотоэлектрическими устройствами. Что же касается бесполезности фотодыхания, то оно может носить приспособительный характер. ИФ являлась предметом длительного естественного отбора, но тем не менее селекционеры должны ее улучшать. Ферменты, структуры и регуляторные процессы, вовлеченные в фотосинтез, могут быть более эффективными подобно другим фундаментальным метаболическим процессам.
1.2.1. Внутривидовые различия по ИФ
Пшеница. Сортовые различия по ИФ флагового листа у пшеницы наиболее заметны при сравнении современных сортов с их дикими сородичами. Преимущество последних, особенно диплоидных форм, по ИФ сочетается с меньшей в сравнении с остальными формами площадью флагового листа, большей скоростью старения и более высоким содержанием N на единицу площади листа. Содержание азота, в свою очередь, положительно коррелирует как с удельной поверхностной плотностью (УПП) листа, так и с ИФ на насыщающем свету (ИФмакс). Более высокая ИФмакс у диплоидов сочетается с меньшим устьичным и остаточным сопротивлением диффузии СО2 и большей плотностью устьиц, что может дать преимущество в аридных условиях. В целом ИФ снижается от диких диплоидов к современным сортам на фоне увеличения размеров клеток мезофилла, числа хлоропластов и активности РБФК в расчете на клетку, за исключением Тritiсит dicoccoides, у которой высокая ИФ сочетается с достаточно крупным флаговым листом. Таким образом, повышение урожайности современных сортов пшеницы в сравнении с дикими предшественниками или сортами ранней селекции не связано с повышением ИФ.
Кукуруза. Селекция на быстрый рост у проростков кукурузы сопровождалась снижением ИФ при одновременном увеличении скорости нарастания площади листьев. Возвратная селекция на высокую урожайность, как правило, не затрагивает ИФ, что показано на 64 инбредных линиях. Некоторым исследователям удалось обнаружить более высокую ИФ у современных гибридов на фоне изменения высоты растений, продолжительности периода до цветения, но урожайность при этом достоверно не увеличивалась.
Соя. Межсортовые различия по ИФ у сои положительно коррелируют с содержанием хлорофилла, азота и числом хлоропластов, но отрицательно — с площадью листьев и урожаем. В сравнительных исследованиях растения, выращенные из крупносемян-
13
ных сортов, как правило, имели более крупные листья, меньшую ИФ, но больший урожай. В то же время ИФ в период интенсивного налива бобов положительно коррелирует с урожаем, что, видимо, отражает влияние аттрагирующей силы акцептора на ИФ. Следует особо подчеркнуть, что ранжирование сортов по ИФ в онтогенезе может меняться. Целенаправленная селекция на высокую ИФ в целом возможна, но урожайность и скорость роста F7-линий с высокой и низкой ИФ — практически не различались.
Горох. ИФ у генотипов гороха варьирует и достаточно хорошо наследуется. Однако когда шесть генотипов гороха, отобранных на высокую или низкую ИФ, сравнивали в полевых условиях, скорость накопления биомассы посева и урожай были выше у генотипов с низкой ИФ в один год исследований, а в другой — разница в ИФ не была связана с индексом листовой поверхности (ИЛП), а относительная скорость роста (ОСР) положительно коррелировала с ИФ.
Картофель. В опытах с 18 клонами картофеля обнаружена высокая положительная корреляция между фотосинтезом листа (ИФ × площадь листа) и урожаем клубней.
Райграс. Различия между генотипами райграса по ИФ обнаружены как при насыщающей фотосинтез освещенности, так и при лимитирующей, причем преимущество по ИФ в различных условиях имеют разные генотипы. Интенсивность фотосинтеза на насыщающем свету отрицательно коррелирует с размерами клеток мезофилла, и селекция на мелкоклеточность приводит к увеличению чистой продуктивности фотосинтеза (ЧПФ), но снижает относительную площадь листьев (ОПЛ), рассчитываемую как отношение площади листьев растения к его массе. В результате относительная скорость роста практически не изменяется. Однако селекция на мелкоклеточность приводит к образованию более крупных семян, давая преимущество в стартовом росте и в конечной надземной биомассе. Генотипические различия по ИФ обнаружены также при высокой концентрации СО2.
Таким образом, по мере селекционного повышения потенциальной урожайности ИФ не увеличивалась. Более того, в большинстве случаев ИФ диких предшественников современных сортов полевых культур оказалась выше, т. е. скорость фотосинтеза непосредственно не лимитировала урожайность и эти параметры коррелировали между собой даже отрицательно.
Если обнаружена положительная корреляция, ИФ определяли в генеративной фазе (рост зерновки или налив бобов), когда существенную роль играла аттрагирующая сила акцептора. Необходимо подчеркнуть, что целенаправленная селекция на высокую ИФ не приводила к повышению урожайности, а в некоторых случаях (кукуруза, горох, люцерна) даже снижала ее. Кроме того, такая селекция может затронуть другие параметры культуры. Например,
14
селекция зерновых на высокие ИФ и УПП листьев часто связана не только с большей толщиной листовой пластинки, но и с большей ее эректоидностью и с меньшим числом более крупных побегов. Итоговый эффект этих изменений зависит от густоты посева и условий выращивания. Наблюдается явный парадокс — у большинства культур имеется значительная вариация по ИФ, которая хорошо наследуется, и поэтому достаточно легко вести селекцию на высокую ИФ. Вместе с тем увеличение ИФ путем повышения освещенности и концентрации СО2 ведет к увеличению урожайности, а генетическое повышение — нет. Этот парадокс был неправильно понят Зелитчем, который предположил, что это игнорирует положительную связь между урожаем и ИФ, определяемой факторами среды. Прояснить этот парадокс можно, лишь доказав в будущем, что селекция на высокую ИФ вносит вклад в увеличение урожайности.
1.2.2. Связь между ИФ и площадью листьев
Как уже отмечалось, ИФ в большинстве случаев (но не всегда) отрицательно коррелирует с площадью листьев, что было продемонстрировано на 16 полевых культурах с коэффициентом корреляции, равным, например, у сои 0,94. Сортовые различия по площади листьев и ИФ у культур не являются независимыми переменными, и их взаимная компенсация представляет главный фактор фотосинтетического парадокса.
Несколько элементов этих компенсаций играют важную роль в экологической адаптации. В пределах культуры доли сухой биомассы и N, приходящиеся на лист, не варьируют существенно в зависимости от генотипа, при условии что растения не сильно различаются по массе и габитусу. Установлена зависимость ОПЛ от УПП для большого числа диких и культурных генотипов риса в широком диапазоне условий выращивания, когда 46 % сухой массы растения сосредоточено в листьях с некоторыми колебаниями этого параметра, определяемого массой растений. Тем не менее следствием большей площади листьев являются меньшая сухая масса и меньшее количество N и соответственно сниженное количество РБФК и других фотосинтетических белков, приходящихся на единицу площади листа в сочетании с пониженными значениями ИФ. Этот феномен известен как «эффект разбавления».
Относительное преимущество любого сочетания ИФ и площади листьев зависит от фазы развития растений, густоты посева и погодных условий. На более ранних фазах развития при низком ИЛП нарастание площади листьев для увеличения поглощения ФАР по меньшей мере компенсирует падение ИФ. В густых же посевах корреляция между площадью листьев и урожаем будет
15
слабее, а с ИФ — сильнее, что было продемонстрировано, в частности, на кормовых бобах. Таким образом, онтогенетическая динамика от крупных листьев с низкой ИФ в ранние фазы онтогенеза к мелким листьям с высокой ИФ при смыкании междурядий в посеве может быть более предпочтительной. Подобная тенденция в некоторой степени наблюдается у зерновых, в частности у пшеницы и кукурузы, в сочетании с изменением положения листьев от горизонтального к вертикальному по мере увеличения площади листьев в онтогенезе растений в посеве. Однако неизвестно, в какой мере это коррелирует с высокой продуктивностью.
Еще одной причиной отбора форм с более крупными листьями, а не с более высокой ИФ является положительная корреляция между размерами листа и плодоэлемента (зерновки). Это хорошо показано на сортах пшеницы и ее диких сородичах. Частично это связано с переходом от диплоидов к гексаплоидам, но в большей степени — от диких форм к диплоидам. Более крупные зерна соответствуют также большей площади листьев и меньшей ИФ у сои и кормовых бобов, где нет изменений в плоидности. Таким образом, установленная закономерность связана не только с плоидностью листьев.
Параллельные изменения размеров клеток листьев могут быть одной из причин подобных аллометрических связей. Однако проекционная площадь клеток мезофилла в процессе селекции у пшениц увеличилась лишь вдвое, слабо коррелируя с площадью поверхности листьев. Масса эндосперма у зерновки при этом практически не изменилась. Наряду с увеличением размеров клеток в результате нарастания плоидности более существенный вклад в увеличение площади листьев и снижение ИФ у пшеницы внесло увеличение числа клеток, приходящееся на единицу площади листа. Отрицательная корреляция между размерами клеток мезофилла и ИФ установлена наряду с пшеницей и у других культур.
Селекция на размеры клеток мезофилла листа у райграса приводила к компенсационным эффектам по площади листьев и ЧПФ. Размеры устьиц и их плотность при этом также варьировали в зависимости от размера клеток. Так, отбор форм с низкой плотностью расположения устьиц с целью снижения водопотребления у ячменя привел к обратным результатам, поскольку параллельно, благодаря более крупным клеткам, увеличивались размеры устьиц, площадь листьев и замедлялось их старение. Таким образом, вследствие комплексной реакции растения селекция на какой-то один показатель затрагивала целую цепь изменений, приводя к противоположным нашему ожиданию результатам. Аналогичная ситуация наблюдалась при селекции на высокую ИФ, приведшей к снижению урожая пшеницы, или селекции на крупность листьев, в результате чего замедлялось их появление (развертывание).
16
Вместе с тем оптимальный размер листа зависит также от уровня освещенности и доступности минерального питания и воды, которые в свою очередь влияют, как и другие факторы регуляции фотосинтеза, на относительный вклад сопротивления прилистного слоя и устьичного сопротивления диффузии СО2.
1.2.3. Связь ИФ с продолжительностью функционирования листа
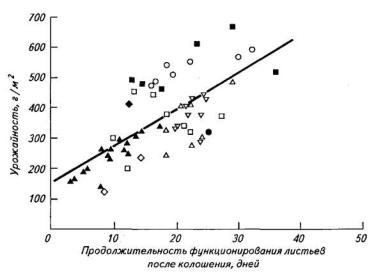
Повышение урожая у современных сортов в сравнении с сортами ранней селекции обусловлено большей продолжительностью фотосинтетической деятельности, количественно характеризуемой фотосинтетическим потенциалом (рис. 4). Отрицательная корреляция между максимальными значениями ИФ и ФП установлена в сравнительных исследованиях диких форм и современных сортов пшеницы, ранне- и позднеспелых сортов сои, но не гибридов кукурузы или генотипов райграса. Однако как у пшеницы, так и у сои более продолжительная фотосинтетическая деятельность форм с низкой ИФ обусловлена более высокой аттрагирующей способностью растущих плодоэлементов. Последняя может оказывать разнообразное влияние на длительность функционирования фотосинтетического аппарата в зависимости
Рис.4. Зависимость зерновой продуктивности пшеницы от продолжительности функционирования листьев после колошения в различных условиях выращивания
(Evans et al., 1915)
17
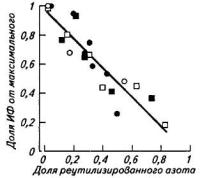
Рис. 5. Связь между скоростью фотосинтеза (% от максимального) и долей реутилизированного из флагового листа азота (символы характеризуют разные уровни азотного питания, заштрихованные — более высокие дозы азотного удобрения)
от относительных скоростей запасания углеводов, жиров или белков в плодоэлементе, а также азотного питания культуры. При доминировании накопления углеводов на фоне благоприятного азотного питания спрос на ассимиляты со стороны акцептора
(зерновки) может продлить фотосинтетическую деятельность листа. Если же происходит интенсивное накопление белков в зерне, а содержание азота в листьях низкое, активизируется реутилизация азота из листьев, в том числе за счет распада белков фотосинтетических ферментов, и листья быстро стареют (рис. 5). Это особенно заметно у высокоурожайных высокобелковых зернобобовых культур. Сортовые различия по продолжительности фотосинтетической деятельности, кроме того, могут объясняться сохранением неодинаковой функциональной активности корневой системы, как, например, у риса в генеративной фазе.
Представляет интерес фотосинтетическая деятельность так называемых ремонтантных гибридов кукурузы с определенной схемой старения листьев: стареют прежде всего листья верхней и нижней трети стебля. Надпочатковые и подпочатковые листья сохраняют фотосинтетическую активность на уровне 20 % от максимальной до полного созревания зерен в початке. Интересно, что подобная схема старения наблюдается также при удалении початка, что свидетельствует о второстепенной роли донорно-акцепторных отношений в проявлении данного феномена. Известны и такие формы, листья у которых в фазе полной спелости початка остаются зелеными и их убирают на силос.
1.2.4. Активность рибулезобифосфат-карбоксилазы/оксигеназы (РБФК/О)
Фермент РБФК/О является главным компонентом белка листа, он составляет 20 % азота и 40 % всего водорастворимого белка у С3-видов. Эти пропорции варьируют от вида к виду и в некоторой мере даже от сорта к сорту. Говоря о межвидовых различиях, необходимо прежде всего упомянуть виды с С3- и С4-типами фотосинтетического метаболизма углерода, причем доля азота листа в РБФК у С4-видов значительно ниже, чем у С3-видов. Среди сор-
18
тов доля водорастворимого белка фермента РБФК колеблется от 48 до 57 % у риса и от 44 до 56 % у люцерны. Обнаружена высокая положительная корреляция между активностью РБФК и максимальной ИФ при световом насыщении у пшеницы, риса и сои.
Лимитирование фотосинтеза РБФК, несмотря на ее значительное количество в листе, в сочетании с высокой молекулярной массой и низким числом оборотов делает ее главным объектом для улучшения. Это объясняется тем, что кинетические свойства фермента представляют анахронизм при нынешних концентрациях СО2 и О2. Однако такая точка зрения является упрощенной, если принять во внимание длительный период интенсивного природного отбора, которому фермент был подвержен. Одним из примеров видимого адаптивного изменения РБФК является различие в кинетических параметрах фермента у С3- и Севидов, причем Км (СО2) ниже у С3-видов, а удельная активность выше у С4-растений.
Внутривидовые различия в активности РБФК менее очевидны. Так, среди 25 сортов риса обнаружены лишь небольшие различия в Км (СО2), VMAKC и удельной активности РБФК, но они более заметны между видами Oryza с геномами А и С. Среди линий ячменя установлены колебания доли белка фермента в общем пуле водорастворимого белка (65—74 %) и удельной активности РБФК. Плоидность также оказывает влияние на вышеуказанные параметры. Так, удельная активность фермента у гексаплоидной пшеницы Т. aestivum in vitro была на 30 % выше, чем у диплоидной Т. топосиссит. Это обусловлено влиянием цитоплазмы генома В на большую субъединицу фермента. Однако указанные различия не сохраняются в системе in vivo.
Сравнительные исследования показали, что характер зависимости ИФ от содержания N в листе, как и ИФ от активности РБФК, у риса и пшеницы одинаков, несмотря на то что количество фермента, приходящееся на единицу площади листа, у риса на 25 % больше (рис. 6). Поэтому теоретически эффективность РБФК у риса можно было бы повысить на 25 %. Однако о регуляторных механизмах, которые могли бы привести к подобным результатам, очень мало известно. Кроме того на трансгенных растениях, выращиваемых при температуре 20 °С, освещенности 300 мкмоль квантов/м2•с, установлено, что содержание белка РБФК у С3-растений необходимо снизить на 40 %, и лишь после этого заметно влияние РБФК на ИФ.
Как показали опыты на трансгенных растениях, в регуляции фотосинтеза существенную роль наряду с РБФК могут играть и другие ферменты цикла Кальвина (фруктозо 1,6-фосфатаза, седугептулезо 1,7-бисфосфатаза, альдолаза и др.). Поэтому попытка представить РБФК как единственный ключевой фермент, определяющий ИФ, может быть ошибочной, особенно если иметь в виду, что как количество, так и активность РБКФ подвержены быстрым изменениям в течение дня в ответ на изменение освещенности и относительно медленным при изменении донорно-акцепторных отношений и концентрации СО2.
19

Рис. 6. Зависимость ИФ флаговых листьев риса и пшеницы от содержания общего азота
(Makino et al., 1988)
1.2.5. Транспорт электронов
Генерация энергии для цикла Кальвина происходит в световой фазе фотосинтеза при фотоиндуцированном транспорте электронов. Увеличение квантового выхода фотосинтеза было бы полезным, но большинство культур, видимо, уже почти достигло максимальной эффективности использования поглощенной радиации. Сравнение 22 контрастных видов полевых культур показало их примерно равные квантовые выходы при разных длинах волн. В опытах с несколькими диплоидными и полиплоидными культурами квантовый выход достиг максимальной величины — 0,08 мольСО2/моль квантов у С3- растений (при пониженной концентрации О2) и 0,052—0,069 — у видов с С4- типом фотосинтеза. Меньший квантовый выход у С4-видов отражает более значительные внутренние энергетические потребности у этой группы видов. Однако при изучении характера зависимости ИФ от поглощенной ФАР при естественной концентрации СО2 и О2 установлена одинаковая величина квантового выхода у С3- и С4-видов, равная 0,05. Видимо, различия в поглощении листом квантов света в полевых условиях оказывают большее влияние на ИФ, чем непосредственно на квантовый выход.
20