

Кошкин
.pdfРис. 69. Этапы формирования элементарного волокна (I):
А — координированный рост волокна; Б, В — стадии интрузивного роста; Г, Д— стадии утолщения клеточной стенки. ТС — точка слома
иклеточной стенки волокна льна (II):
А— первичная клеточная стенка (ПКС) и срединная пластинка (СП) тшш, Б— начало формирования вторичной клеточной стенки (ВКС), В— образование двух слоев вторичной клеточной стенки (I — ^ш, II — i=i), У—Ж—последующие стадии созревания —
уменьшение толщины слоя I. А—Д— период быстрого роста. На примере волокна из средней части стебля показано его дальнейшее развитие в фазе зеленой (Е) и желтой (Ж) спелости
6.4.1. Дифференциация волокон
Волокна льна по происхождению являются первичными, образующимися в конусе нарастания стебля из первичной меристемы. Лубяные волокна льна образуются из прокамбия и составляют элементы протофлоэмы. В ходе развития ситовидные трубки и клетки-спутники первичной флоэмы разрушаются, а клетки волокон становятся шире и длиннее. Когда следы проводящих элементов исчезают, остатки протофлоэмы представляют собой однородную ткань, состоящую из одного типа клеток — молодых волокон. На продольных срезах молодые волокна можно идентифицировать на расстоянии 0,5 мм от апекса на уровне 4—5-го примордия.
221
6.4.2.Удлинение волокон
Вначале своего развития волокна, находящиеся в апикальной области стебля (несколько верхних миллиметров), растут скоординированно со всеми окружающими тканями. Такой тип роста называют координированным. Межклеточные контакты при этом не нарушаются, и волокна связаны с соседними клетками плазмодесмами. На стадии координированного роста волокна становятся длиннее, чем клетки окружающих тканей, за счет более раннего прекращения делений. Для этой стадии роста волокон характерны вытянутая прямоугольная форма и тупые закругленные концы. На стадии координированного роста клетки лубяных волокон не достигают большой (100—200 мкм) длины и имеют диаметр 4—7 мкм.
Вдальнейшем большинство тканей стебля перестает расти, а лубяные волокна продолжают удлиняться, внедряясь концами между соседними клетками. Такой тип роста называют интрузивным. Наряду с удлинением волокна увеличиваются в диаметре с 7 до 15—25 мкм. Для лучшего проникновения между клетками их кончики заостряются и из-за механического сопротивления окружающих тканей часто становятся искривленными. Клетки волокон растут всей поверхностью (поверхностный, или диффузный, рост). При этом неизбежно нарушаются контакты между клетками и разрушаются плазмодесмы.
Удлинение всех волокон завершается выше точки слома. Ниже нее происходят только утолщение клеточной стенки и ее модификация, приводящая к созреванию волокна. Зная, что скорость роста растений в период быстрого роста составляет несколько сантиметров в день, можно рассчитать продолжительность различных стадий роста лубяного волокна. Координированный рост занимает несколько часов, интрузивный — около двух дней, а утолщение и созревание клеточной стенки — до окончания вегетации, т. е. почти два месяца. Хотя основная масса волокна формируется на стадии утолщения клеточной стенки, скорость процесса образования компонентов клеточной стенки высока и в период удлинения.
6.4.3.Утолщение клеточной стенки и созревание волокон
Формирование вторичной клеточной стенки в некоторых волокнах начинается уже выше точки слома, охватывая в первую очередь волокна, расположенные в наружной части пучка (рис. 70). Оно происходит неравномерно по длине волокна, начиная с его средней части.
На электронно-микроскопических фотографиях видно, что вторичная клеточная стенка в начале своего формирования имеет характерную полосатую структуру, где темные полосы чередуются
222
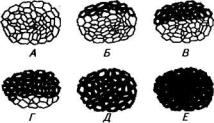
Рис. 70. Последовательные стадии утолщения клеточных стенок в пучке лубяных волокон (пояснения даны на рис. 69)
со светлыми участками. Позднее вторичная клеточная стенка выглядит двухслойной: в наружной части образуется светлый слой, имеющий в отличие от полосатого внутреннего тонкофибриллярную структуру. Именно возникновение такого слоя вторичной клеточной стенки и определяет изменения механических свойств стебля в точке слома. По мере созревания волокна внутренний слой постепенно уменьшается и в зрелом волокне исчезает. Такое изменение структуры вторичной клеточной стенки может быть связано с постсинтетическими изменениями составляющих ее полимеров (характера связей, степени гидратации), приводящими к уплотнению клеточной стенки в процессе созревания волокна.
Структура клеточной стенки волокон, находящихся в разных участках стебля растения в период быстрого роста (А—Д), изображена на рисунке 69 II. На примере волокна из средней части стебля показана дальнейшая модификация клеточной стенки (Е, Ж), которую последовательно претерпевают все волокна на следующих фазах развития растений.
Если во время своего роста волокна увеличиваются и в толщину (увеличивается объем цитоплазмы и вакуоли), то с началом образования вторичной клеточной стенки увеличение диаметра волокна незначительно. Утолщение клеточной стенки неизбежно приводит к сокращению объема протопласта: в конце развития почти весь люмен клетки заполняется клеточной стенкой. Уменьшение объема протопласта происходит частично за счет постепенной деградации цитоплазмы, но в основном благодаря уменьшению величины центральной вакуоли. Излишек мембран, образующийся в результате этого процесса, а также экзоцитоза полисахаридов матрикса клеточной стенки скапливается в периплазматическом пространстве в виде пузырьков и трубочек. Извилистый контур плазмалеммы и наличие мембранных структур в периплазме — характерные особенности ультраструктуры волокон льна. Излишки тонопласта могут накапливаться внутри вакуоли.
Й период утолщения клеточной стенки цитоплазма в волокне распределена неравномерно: на одних участках она имеет вид узкого слоя между стенкой и центральной вакуолью, которая занимает основную часть полости клетки, на других участках, в том числе в кончиках, цитоплазма образует скопления со многими органеллами и центральная вакуоль отсутствует. В целом волок-
223
на более насыщены цитоплазмой, чем смежная коровая паренхима. Протопласт в волокне сохраняется долгое время вплоть до фазы желтой спелости. Однако цитоплазма расположена тонким пристенным слоем и имеет признаки деградации — аппарат Гольджи теряет активность, происходит просветление матрикса митохондрий, наблюдаются мультивезикулярные тела в вакуоли и периплазме. К фазе полной спелости люмен волокна сильно сужается, и протопласт отмирает. Механизмы и регуляция этого процесса, во многом определяющего наличие цитоплазматических «примесей» в зрелом волокне, у льна практически не изучены.
6.4.4. Значение периода быстрого роста растений для формирования волокна
Ключевым для формирования лубяных волокон этапом в развитии растений льна является период быстрого роста. В это время на растущем стебле льна можно найти волокна, находящиеся на всех основных этапах формирования (см. рис. 69, I). Дифференциация и удлинение всех клеток лубяных волокон заканчиваются в период быстрого роста. Поэтому все характеристики качества урожая, которые зависят от количества клеток волокон и их длины, определяются уже на стадии быстрого роста и в дальнейшем не могут быть изменены. На более поздних этапах развития апикальная меристема перестраивается на формирование соцветий, точка слома исчезает, в клетках волокон происходят лишь процессы утолщения клеточных стенок и созревания волокон.
6.5. Состав льняного волокна
Основную массу (85—87 %) льняного волокна составляют полисахариды клеточной стенки. Полисахариды — полимеры, состоящие из различных моносахаридов. Наиболее распространенными мономерами в полисахаридах растительной клеточной стенки являются глюкоза, галактоза, манноза, рамноза, фруктоза, которые содержат 6 атомов углерода, а арабиноза и ксилоза — 5 атомов углерода. В отличие от нуклеиновых кислот и белков, где все мономеры связаны между собой одним и тем же типом связи с участием одних и тех же атомов, каждый моносахарид способен образовать связь с участием разных атомов углерода. Разнообразие в типах мономеров и в положениях, по которым осуществляется связь между ними, приводит к необоз-
224
римому множеству полисахаридных структур. Характеризуя полисахарид, необходимо указать: 1) из каких мономеров он состоит; 2) D- или L-форма сахара; 3) по каким номерам атомов углерода осуществляется связь; 4) тип связи (а- или р-); 5) форму молекулы (линейная или разветвленная); 6) степень полимеризации. Степень полимеризации полисахаридов клеточной стенки может различаться у разных молекул одного и того же полимера. Современные модели клеточной стенки высших растений изображают ее как сложную многокомпонентную систему, основу которой составляют три типа полисахаридов: целлюлоза; поперечно-связывающие гликаны (раньше их называли гемицеллюлозы); пектиновые вещества. Их соотношение в зрелом льняном волокне составляет 75 : 10: 15. Состав волокна существенно изменяется в ходе его формирования, например, в молодых клетках содержание целлюлозы значительно ниже (30 %).
6.5.1. Целлюлоза
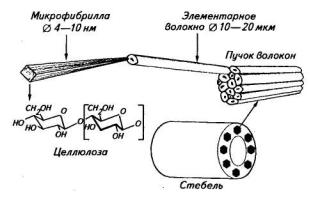
Основной компонент льняного волокна — целлюлоза, молекулы которой устроены, казалось бы, простейшим способом. Они представляют собой линейные цепочки гомополимера, т. е. полимера, состоящего из мономеров одного типа. Мономером целлюлозы является p-D-глюкоза. Все остатки глюкозы соединены между собой одним типом связи с участием первого атома углерода одной молекулы глюкозы и четвертого атома углерода следующей молекулы. Таким образом, химическое название целлюлозы Р-1,4-Б-глюкан. Степень полимеризации молекул целлюлозы в льняном волокне составляет около 10 тыс.
Рис. 71. Уровни организации льняного волокна
225
Молекулы целлюлозы существуют в клеточной стенке не каждая сама по себе, а в виде надмолекулярных образований, называемых микрофибриллами. Микрофибрилла (диаметр 10 нм) состоит в поперечнике из нескольких десятков молекул целлюлозы. Внутри микрофибриллы молекулы целлюлозы начинаются и кончаются в разных местах, поэтому микрофибрилла в целом имеет большую длину. Микрофибрилла образуется за счет многочисленных водородных связей между молекулами целлюлозы. Многочисленные микрофибриллы целлюлозы образуют в клеточной стенке слои. Характерной чертой целлюлозы льняного волокна является высокая упорядоченность ее структуры с образованием кристаллических участков (рис. 71).
6.5.2. Нецеллюлозные полисахариды
Нецеллюлозные полисахариды в значительной степени определяют специфику свойств льняного волокна и отличают его, например, от хлопкового аналога, который практически полностью состоит из целлюлозы. Они играют важную роль в детерминации водопоглотительной способности, набухания, эластичности. Номенклатура этих полимеров построена так, что последним в слове называются моносахариды, из которых сформирован остов молекул, а перед ним ставят названия тех моносахаридов, которые встроены в боковые цепочки. Простой перечень идентифицированных полисахаридов клеточной стенки показывает, насколько сложно она устроена. В клеточной стенке льняных волокон обнаружены ксилоглюкан, глюкуроноксилан, глюко(галакто)маннан, арабиногалактан, арабинан, полигалактуроновая кислота, рамногалактуронан и др. (табл. 38). На этапе формирования волокон в них присутствует особый полисахарид — высокомолекулярный га-лактан. Не исключено, что будут идентифицированы и другие полисахариды.
Нецеллюлозные полисахариды клеточной стенки разделяют на две группы. К первой группе относят нейтральные полисахариды, большинство из которых связывают между собой микрофибриллы целлюлозы (так называемые поперечно связывающие гли-каны). Вторая группа включает полисахариды, содержащие модифицированные сахара — уроновые кислоты, которые придают полимеру отрицательный заряд; их называют пектиновые вещества. Деление на группы общепринято, но достаточно условно, поскольку некоторые полисахариды, традиционно относимые к нейтральным, содержат небольшое количество уроновых кислот (например, глюкуроноксилан). Вместе с тем пектиновые вещества иногда содержат нейтральные боковые цепочки, многократно превышающие по массе отрицательно заряженный остов молекулы.
226
38. Нецеллюлозные полисахариды волокон льна
|
Моносахариды |
Моносахариды |
|
|
Полисахарид |
остова, тип связи |
боковых цепочек, |
Примечания |
|
между ними, место |
тип связи между |
|||
|
|
|||
|
ветвления |
ними |
|
|
Ксилоглюкан |
D-глюкоза, β-1,4, |
D-ксилоза, β-1,2 |
Кси3Глю4 |
|
ветвление С-6 |
||||
|
|
|
||
|
D-ксилоза, β-1,4, |
Одиночная D- |
|
|
Ксилан |
глюкуроновая |
|
||
ветвление С-6 |
|
|||
|
кислота |
|
||
|
|
|
||
|
L-рамноза, |
|
|
|
|
D-галактуроновая |
|
|
|
Рамногалактуронан I |
кислота, звенья (4-α- |
— |
— |
|
ГалУК-1,2-α-Рам-1), |
||||
|
|
|
||
|
ветвление С-4 |
|
|
|
|
рамнозы |
|
|
|
Арабиногалактан |
D-галактоза, β-1,3, |
L-арабиноза |
— |
|
ветвление С-6 |
||||
|
|
|
||
Полигалактуроновая |
D-галактуроновая |
— |
— |
|
кислота |
кислота, α-1,2 |
|||
|
|
|||
|
|
|
Может являться |
|
Арабинан |
L-арабиноза, α-1,5 |
L-арабиноза |
боковой цепочкой |
|
|
|
|
рамногалак-туронана |
|
Глюко(галакто)-маннан, |
D-манноза, Р-1,4 |
D-глюкоза, |
— |
|
D-галактоза |
||||
|
|
|
||
|
|
|
Степень |
|
|
|
|
полимеризации > 10 |
|
|
D-галактоза, β-1,4, |
Одиночная |
тыс., водорастворим, |
|
Галактан |
присутствует только в |
|||
ветвление С-2 или С-3 |
D-галактоза |
клетках волокон и |
||
|
||||
|
|
|
только на |
|
|
|
|
определенной стадии |
|
|
|
|
их развития |
Остов молекул поперечно связывающих гликанов устроен по тому же принципу, что и молекулы целлюлозы. Но к этому остову с определенной периодичностью добавлены боковые ответвления, которые нарушают линейность молекулы полисахарида. Другим способом получения изгибов молекулы является чередование типов связи мономеров внутри нее. В результате получается изогнутая во многих местах молекула, содержащая линейные участки. Этими линейными участками молекулы поперечно связывающих гликанов взаимодействуют как с молекулами целлюлозы, так и между собой. В результате формируется прочная сеть, состоящая из двух типов молекул: целлюлозы и поперечно связывающих гликанов.
В роли поперечно связывающих гликанов у разных организмов выступают различные полисахариды. У двудольных основным
227
поперечно связывающим гликаном является ксилоглюкан, т. е. полисахарид, остов которого построен из глюкозы, а боковые цепочки — из ксилозы. Этот полимер содержится и в волокнах льна. Наряду с ним функции поперечно связывающих гликанов в волокне выполняют, вероятно, глюкуроноксилан и глюко(галакто)-маннан.
Пектиновые вещества — полисахариды, образованные остатками главным образом галактуроновой кислоты. Полигалактуроновая кислота — гомополимер из остатков галактуроновой кислоты, связанных а-1,4-связью. Во многом благодаря наличию полигалактуроновой кислоты (в отличие от нейтральных полисахаридов) клеточная стенка имеет отрицательный заряд и ведет себя как нерастворимый полианион.
Рамногалактуронан I — гетерополимер, остов которого состоит из рамнозы и галактуроновой кислоты. К остаткам рамнозы могут быть присоединены различной длины цепочки арабиногалактанов, арабинанов и галактанов. Типы боковых цепочек рамногалактуронана в значительной степени определяют пористость клеточной стенки, ее заряд, рН и баланс ионов.
В льняном волокне присутствуют и полигалактуроновая кислота, и рамногалактуронан I. Они локализованы главным образом в срединной пластинке. Однако часть (40 % уроновых кислот) пектиновых веществ волокна очень прочно связана с целлюлозой, а значит расположена в глубинных слоях клеточной стенки. Поэтому в технологиях, где необходима чистая целлюлоза с ненарушенной структурой, льняное волокно вряд ли пригодно.
Количественно оценить содержание различных полисахаридов в клеточной стенке непросто. Это связано со сложностями разделения индивидуальных полисахаридов и характеристики разнообразных связей между мономерами. Традиционным приемом является гидролиз полисахаридов до мономеров и определение соотношения между ними. Моносахаридный состав полисахаридов клеточной стенки зрелых волокон льна (до мочки) следующий (моль %): рамноза — 0,2, арабиноза — 0,6, галактоза — 3,9, глюкоза — 83,2, ксилоза — 1,9, манноза — 4,8, уроновые кислоты — 5,4. Необходимо помнить, что один и тот же моносахарид может входить в состав нескольких разных полисахаридов. Например, в волокнах льна глюкоза входит в состав целлюлозы и ксилоглюкана, ксилоза — в состав ксилоглюкана и глюкуроноксилана. Методы, позволяющие установить типы связей между мономерами и идентифицировать полисахарид, к сожалению, не могут служить основой для количественной оценки, поэтому содержание индивидуальных полисахаридов в клеточной стенке волокон льна остается невыясненным, как, впрочем, у большинства других растительных объектов.
228
Еще сложнее определить взаиморасположение различных полисахаридов в клеточной стенке и их локализацию в различных слоях. Основной используемый при этом подход — иммуноцитохимия. Есть и другие, более косвенные подходы. По результатам обработки волокна экзогенными ферментами, гидролизующими полисахариды, можно сделать вывод, что ксилоглюкан и глюкуроноксилан расположены в глубине клеточной стенки и недоступны для воздействия индивидуальных экзогенных ферментов.
Важнейшее свойство клеточной стенки — ее мозаичность, т. е. неоднородность различных участков, составляющая основу для четко локализованных реакций в клеточной стенке. Мозаичность клеточной стенки хорошо выявляется при использовании иммунохимических методов окрашивания, без них ее трудно обнаружить. Особые свойства участков клеточной стенки определяются иногда нюансами в строении полимеров, например степенью метилирования уроновых кислот в пектиновых веществах. Известно, что в льняном волокне мозаично расположены специальные «довески» на некоторых полисахаридных цепочках — молекулы феруловой кислоты. Характеристика мозаичности клеточной стенки льняных волокон, безусловно, требует дальнейших исследований, тем более что она может служить основой многих процессов, наблюдаемых при переработке льна.
Особый полисахарид клеточной стенки волокон льна — высокомолекулярный β-1,4-D-галактан. Этот полимер примечателен тем, что при степени полимеризации, сравнимой с таковой у целлюлозы, он является водорастворимым соединением. Однако еще больший интерес вызывает то, что, во-первых, это тканеспецифичный полимер и в стебле льна содержится только в клетках лубяных волокон, а во-вторых, он присутствует только на определенной стадии развития волокна. Синтез тканеспецифичного галактана начинается в точке слома; выше нее (т. е. в удлиняющихся волокнах) этот полимер не обнаружен. Судя по динамике и локализации синтеза, тканеспецифичный галактан принимает участие в процессах созревания клеточной стенки волокна. Он появляется в тот момент, когда начинается формирование особой структуры клеточной стенки, которая на электронномикроскопических снимках выглядит как тонкофибриллярная. В дальнейшем галактан подвергается модификации и (или) гидролизу и в зрелом волокне не обнаруживается. В зрелом волокне присутствуют полисахариды, которые могут быть остатками этого высокомолекулярного полимера. Разработка подходов к изменению метаболизма тканеспецифичного галактана исключительно важна как для понимания его функции и механизма действия, так и для модификации качества льняного волокна.
229
6.5.3. Лигнин и другие фенольные соединения
Клеточные стенки высших растений содержат, помимо сложных полисахаридов и белков, фенольные соединения, которые обычно подразделяют на лигнин и оксикоричные кислоты. Содержание этих соединений особенно велико в древесных растениях, где оно достигает 50 % от сухой массы клеточных стенок. Согласно общим представлениям, лигнин — это гетерополимер, образующийся из трех основных мономеров: n-кумарового, кониферилового и синапового спиртов, называемых монолигнолами. Он является одним из самых стабильных органических соединений и трудно поддается экстракции из растительных тканей. Монолигнолы могут быть соединены между собой в молекуле лигнина разнообразными эфирными и углеродуглеродными связями. Оксикоричные кислоты, в основном феруловая и n-кумаровая, образуют эфирные связи с различными полимерами клеточных стенок, придавая клеточной стенке дополнительную прочность.
В волокнах льна содержание лигнина очень низкое — 0,5 % и менее. Ранее использовавшиеся методы давали завышенную оценку (2—5 %). Вторичная клеточная стенка льняного волокна не лигнифицируется. В конце вегетации лигнин откладывается в срединных пластинках волокон и углах клеток, при этом лигнификация затрагивает (неизвестно, по какому принципу) лишь часть волокон. При излишней лигнификации качество волокна снижается, что является одной из причин сбора урожая льна на волокно до полного созревания семян.
Содержание еще одного типа фенольных соединений клеточной стенки
— оксикоричных кислот (преимущественно феруловой кислоты) еще меньше
—около 0,1 %. Однако даже в такой низкой концентрации и лигнин, и оксикоричные кислоты существенно влияют на свойства волокна.
6.5.4. Липиды, воск и другие органические соединения
Содержание различных липидных соединений и воска в льняном волокне около 1,5 %. Эти вещества существенно влияют на органолептические свойства волокон. Около 1 % массы клеточной стенки составляют белки, которые пока слабо охарактеризованы. Среди них присутствуют как структурные белки клеточной стенки, так и ферменты, участвующие в ее метаболизме. Экстрактивные низкомолекулярные вещества, среди которых различные флавоноиды, гликозиды, органические кислоты, жирные кислоты и др., составляют 2—4 % массы волокна.
230