

Кошкин
.pdfлокна лучших кряжевых льнов был выше, чем местных беспородных, на 53 %, а урожай семян — на 12 %. Кряжевые льны имели большое значение как исходный материал для селекции.
Селекция льна-долгунца в России стала развиваться с начала XX в. На разных этапах селекции менялись ее методы, решались различные проблемы, для отбора использовались признаки, соответствовавшие пониманию физиологии формирования урожая в данный период.
Основным методом селекции при создании первых сортов был индивидуальный отбор высокорослых растений (с учетом комплекса внешних признаков) в разреженном луночном посеве. Однако корреляционная связь между косвенными признаками и содержанием волокна была весьма незначительна. Практика селекционной работы показала, что образцы льна, сходные по морфологическим признакам, часто различались как по абсолютному, так и по относительному содержанию волокна в стеблях. Таким образом, селекция, основанная на использовании морфологических признаков, была бесперспективной и ненадежной. Кроме того, изучение визуальных морфологических признаков вследствие их сильной изменчивости требовало больших затрат времени и труда.
Широкая селекционная работа с льном-долгунцом была развернута в связи с созданием в 1930 г. Всесоюзного научно-исследовательского института льна с сетью зональных льняных опытных станций, полей и опорных пунктов.
Врезультате селекционной работы в 1930—1933 гг. были районированы первые сорта, которые значительно превосходили по урожаю волокна местные сорта льна. Вместе с тем новые сорта имели и существенные недостатки: значительно поражались болезнями, особенно ржавчиной; характеризовались низким урожаем семян, поскольку отбор вели в соответствии с представлением
отом, что малокоробочность связана с высокой волокнистостью. В процессе дальнейших исследований выяснилось, что корреляция между малокоробочностью и высокой волокнистостью незначительна.
С 1930 г. в основу методики селекции льна-долгунца были положены метод определения содержания волокна в единичных стеблях с помощью варки в щелочи и метод выращивания льна в ящиках в вегетационном домике для сравнительных испытаний, оценки и отбора в возможно более выравненных условиях. Эти методы совершенствовались и применялись в селекционной практике до 1949 г. В 30-е гг. XX в. для оценки устойчивости льна к болезням (фузариоз и ржавчина) стали применять провокационные посевы.
Вкороткие сроки (с 1934 по 1936 г.) путем индивидуального отбора из русских кряжевых льнов (главным образом из Печорского
261
и Зарецкого кряжей) были выведены и районированы более ценные высоковолокнистые сорта (табл. 52). Селекционные сорта, переданные в производство с 1934 по 1936 г., характеризовались устойчивостью к ржавчине. Считалось, что проблема выведения и внедрения в производство сортов льнадолгунца, устойчивых к ржавчине, решена, но в 1943 г. обнаружили значительное поражение ржавчиной сортов, ранее устойчивых к этой болезни. В последующие годы ржавчиной болели все ранее устойчивые к ней сорта. Причиной описанного явления стало возникновение новой расы (или рас) ржавчины, приспособившейся к жизни на сортах льна, прежде устойчивых к ней, и вновь возник актуальный и поныне вопрос об изучении механизмов устойчивости.
Большинство сортов, выведенных в 1933—1934 гг., были склонны к полеганию. В России на устойчивость сортов льна к полеганию обращалось мало внимания, так как она не имела большого хозяйственного значения. В 1933—1934 гг. стали активно внедрять механизированную уборку льна. Полегание могло нанести льноводству довольно ощутимый урон, так как полегший лен нельзя было убрать существовавшими в то время теребилками. Устойчивость к полеганию стала одним из приоритетных направлений в селекции.
Если в первые годы при отборе лучших форм из кряжевых льнов ящичные посевы служили своего рода «ситом» (так как кряжевые льны являются популяциями с консервативной наследственностью), то при работе с гибридным материалом использование ящичных посевов стало помехой в селекционном процессе, а выведенные сорта оказались непригодными для использования на полях. Несостоятельность методов стала понятной в 1949— 1951 гг., когда из шести первых гибридных сортов три были сняты с районирования. В результате усовершенствования методов селекции и переноса селекционных посевов из вегетационного домика в полевые условия, а также в связи с использованием тепловой мочки для выделения волокна из стеблей появилась возможность оценивать селекционные номера не только по количеству, но и по качеству волокна.
Это позволило ускорить темпы селекции и передать в государственное сортоиспытание сорта, отличающиеся более полным комплексом хозяйственно полезных признаков. В 1960—1967 гг. районированы сорта, выведенные методом гибридизации, которые имели более высокий урожай волокна и его содержание в стеблях (см. табл. 52). Появился ряд сортов, устойчивых к полеганию.
В последние десятилетия дополнительным требованием к селекции стало эффективное использование культурами минеральных удобрений. К сожалению, изучение сортовой отзывчивости генотипов различных сельскохозяйственных растений и ее физиологической природы почти не коснулось культуры льна. Тем не менее
262
52. Хозяйственные характеристики сортов льна на разных этапах селекции
Период |
Сорта |
Урожайность, ц/га |
Содержание |
Устойчивость |
|||
селекции |
соломки |
волокна |
волокна, % |
к полеганию |
к болезням |
||
|
|||||||
Ранняя |
Местный лен (народной селекции) |
32 |
4 |
|
Не определяли |
||
селекция |
Кряжевые льны |
39 |
5 |
20-21 |
Не определяли |
||
1930— |
823/3, 806/3, Псковский улучшенный, |
|
|
|
|
Неустойчивы и |
|
1933 гг. |
0107, 0109, 0113, 0120, 0262, 0264, 0266 |
38-54 |
5-10 |
18-20 |
Неустойчивы |
устойчивы |
|
|
Альфа, ДСЗО, ДСЗЗ, 2052, Д83 |
|
|
|
|
|
|
1934— |
1288/12, Прядильщик, Светоч, 1908, |
|
|
|
|
Устойчивы и |
|
1936 гг. |
Ударник, Победитель, Текстильщик, |
50-60 |
8-11 |
20-26 |
Неустойчивы |
неустойчивы |
|
|
Стахановец, Омега |
|
|
|
|
|
|
1946- |
И5, И7, И9, И2, А2176, Л1120, |
48-61 |
10-13 |
17-21 |
Неустойчивы |
Неустойчивы и |
|
1951гг. |
Вайжгантас |
и устойчивы |
среднеустойчивы |
||||
|
|
|
|||||
1960— |
Псковский 1, И16, Спартак, Т5, Т7, Т9, |
59-65 |
10-15 |
20-27 |
Среднеустой- |
Неустойчивы и |
|
Т10, ЛД147, Шокинский, ВНИИЛ2, |
|||||||
1967 гг. |
чивы |
среднеустойчивы |
|||||
|
ВНИИЛЗ, ВНИИЛ8, П255 |
|
|
|
|
|
|
1968— |
Богатырь, Рекорд, Старт, Мир, Космос, |
58-68 |
9-12 |
20-24 |
Устойчивы |
Среднеустойчивы |
|
1971 гг. |
Конкурент, Маяк, Смена |
и неустойчивы |
|||||
1985— |
Новоторжский, Призыв 81, С108, |
40-68 |
9-20 |
18-30 |
Устойчивы |
Неустойчивы, |
|
1990 гг. |
Славный 82, Мрия, Лазурный, Союз, |
среднеустойчивы |
|||||
Сальдо, Дашковский, Томский 16 |
|
|
|
|
и устойчивы |
||
|
А29, Алексим, А93, Ленок, Могилевский |
|
|
|
|
|
|
1993— |
2, Томский 17, Томский 18, Нептун, |
40-68 |
11-18 |
24-30 |
Устойчивы |
Среднеустойчивы |
|
2000 гг. |
Кром, Русич, Восход, Синичка, Белочка, |
и устойчивы |
|||||
|
Смолич |
|
|
|
|
|
|
известно, что его различные линии и сорта, безусловно, имеют разную отзывчивость на уровень минерального питания, причем она определяется не мощностью ассимиляционного аппарата и корневой системы и не абсолютной величиной поглотительной способности корневой системы, а направленностью и амплитудой изменения этих показателей по сравнению с неудобренными растениями.
К 1985 г. окончательно определились задачи и направления селекции льна. Современному производству необходимы сорта льна со следующими качествами:
среднеспелые и раннеспелые; высокопродуктивные:
урожайность волокна 12—18 ц/га; содержание волокна 26—30 %; качество длинного волокна — номер 15—16; урожайность семян 7-11 ц/га;
интенсивного типа, т. е. пригодные для возделывания по индустриальной технологии;
отзывчивые на минеральные удобрения; устойчивые к полеганию и болезням;
сдружным созреванием семян;
сустойчивой урожайностью.
Селекционеры работают и в менее традиционных направлениях, например, выводят сорта озимого льна, специализированные «рекультивационные», обеспечивающие очищение почвы от тяжелых металлов, а также декоративные сорта.
Таким образом, на протяжении многих лет новые селекционные сорта льна получали эмпирически. Сначала селекцию проводили по косвенным морфологическим признакам, которые далеко не всегда коррелировали с величиной урожая, его выходом и качеством. Позднее для селекции стали использовать другие (прямые хозяйственные) признаки: урожай и выход волокна, семян, устойчивость к полеганию, болезням, еще позднее — отзывчивость различных сортов на уровень минерального питания.
Однако более перспективны показатели, характеризующие продукционный процесс: фотосинтетический потенциал, содержание хлорофилла, чистая продуктивность фотосинтеза, интенсивность фотосинтеза, дыхания, т. е. интенсивные показатели. При их изучении необходимо установить корреляционные связи этих показателей с урожаем и выходом волокна. К примеру, новые высокопродуктивные позднеспелые сорта и образцы льна характеризуются более высоким содержанием хлорофилла и продолжительной продуктивной фотосинтетической деятельностью (сохраняют высокие значения ЧПФ на поздних этапах онтогенеза). При этом величина ЧПФ положительно коррелирует с выходом волокна (+0,79), площадь листьев имеет положительную корреляционную связь с урожаем соломы (+0,8), но отрицательную с выходом волокна (—0,52). Однако таких полных данных немного.
Выведение высококачественных сортов затруднено тем, что до сих пор неясны механизмы регуляции качества волокна. В то же время целенаправленная селекция на отдельные параметры качества практически не велась, что объясняется отсутствием экспресс-методов их оценки, особенно на первых этапах селекции. Более того, метод гибридизации, применяемый в селекции льна-долгунца с 30-х гг. прошлого века без надежных методов отбора по признаку качества, вероятно, привел к разрушению коадаптированных блоков генов «качества волокна», созданных за столетия народной селекции методом индивидуального отбора. В настоящее время четко проявляется тенденция снижения показателей качества чесаного волокна (гибкость, тонина) у сортов, выведенных методами простой и особенно сложной гибридизации, по сравнению с сортами, созданными методами индивидуального отбора из кряжевых форм льна (таких как Светоч, 806/3, которые относятся к первой группе по прядильным свойствам).
Современные сорта мало различаются по качеству, так как при их выведении местные и кряжевые формы использовались незначительно. Таким образом, необходим поиск генетических источников отдельных параметров качества в коллекции местных и кряжевых форм льна. Основным источником генетического разнообразия стали коллекция льна ВИР и Национальная коллекция русского льна ВНИИ льна, насчитывающие соответственно более 5700 и 6000 образцов. Результаты испытания сортов (1993— 2000 гг.), включенных в Госреестр селекционных достижений, допущенных к использованию и давших наивысшие показатели урожая волокна хорошего качества и семян, представлены в таблице 52.
6.17. Биотехнология льна
Достижения биотехнологии, которые для многих других культур, например для хлопка, уже давно выражаются в доведении до выращивания трансгенных растений на полях, для льна пока не реализованы. Лен-долгунец
— сложный для культивирования in vitro вид, для которого до сих пор не разработаны условия массовой регенерации растений. Масличный лен более пригоден для биотехнологических подходов, но и для него получение новых форм находится пока еще на стадии модельных опытов. Тем не менее, оценивая развитие современных технологий на других видах растений, можно утверждать, что в ближайшем будущем они будут разработаны и для льна. Основным будет вопрос, какие именно гены нужно изменить или добавить для достижения желаемого эффекта. Поэтому характеристика формирования волокон в терминах молекулярной генетики становится особенно актуальной задачей. Можно ожидать, что первыми возникнут новые формы,
265
обладающие устойчивостью к определенным патогенам, поскольку такая устойчивость может обеспечиваться небольшим или даже одним числом генов. Перспективна модификация генов, задействованных в формировании полисахаридов клеточной стенки, особенно в сочетании с тканеспецифичными промоторами, но идентификация и тех и других требует дальнейших исследований.
Контрольные вопросы и задания
1. Почему лубяные волокна льна считаются первичными? Где расположены лубяные волокна в стебле льна и в состав какой ткани они входят? 2. В чем различие ботанического и технического определения волокна? 3. Какие стадии формирования волокон характерны для каждой из фаз развития растений? Какие процессы определяют изменение механических свойств стебля в точке слома? 4. В чем отличие координированного и интрузивного роста волокон? 5. Охарактеризуйте группы полисахаридов, входящие в состав клеточной стенки лубяного волокна. 6. Какие процессы являются источниками сахаров в лубяных волокнах? Как изменяются донорно-акцепторные отношения в ходе развития растения? 7. Какие анатомические и биохимические характеристики влияют на качество льняного волокна? 8. Какие характеристики стебля льна определяют устойчивость к полеганию? 9. Назовите критические периоды в потребности питательных веществ у растений льна. 10. Каковы особенности влияния основных элементов минерального питания (N, Р и К) на фотосинтетическую деятельность и продуктивность льна-долгунца? 11. Каковы требования растений льна к освещенности и способы ее регуляции в посевах льна? 12. Какой из показателей фотосинтетической деятельности растений льна в наибольшей степени определяет величину урожая? 13. В какой период роста растение льна наиболее чувствительно к засухе и с чем это связано? 14. Назовите основные этапы селекции льнадолгунца и применяемые критерии отбора.
266
Глава 7 САХАРНАЯ СВЕКЛА
Сахарная свекла (Beta vulgaris L.) принадлежит к роду Beta L., семейству маревых (Chenopodiaceae). В Западной Европе и России она возникла в конце XVIII — начале XIX в. в результате скрещивания корнеплодных форм с листовыми. В то время сахаристость корнеплодов составляла всего 7—10 % при низком содержании в них сухого вещества, однако уже в начале XX в. были выведены сорта с сахаристостью корнеплодов до 18 %. В мировом производстве сахара в последние десятилетия на долю сахарной свеклы приходится около 30 %. Определенную практическую ценность представляют также отходы свеклосахарного производства — патока (меласса), жом, дефекат.
Со второй половины XX в. достигнут значительный прогресс в создании высокопродуктивных сортов и гибридов сахарной свеклы. Разработаны интенсивные технологии возделывания этой культуры с использованием преимущественно семян гибридов. В странах Западной Европы средняя урожайность корнеплодов сахарной свеклы в последние десятилетия составляет 50—60 т/га при содержании сахара 18 %. В Российской Федерации эти показатели в 2002 г. составили соответственно 21,8 т/га и 13,4 %.
В селекции сахарной свеклы существуют три направления: урожайное, сахаристое и урожайно-сахаристое. Сорта и гибриды урожайного направления отличаются от остальных наибольшей массой корнеплодов, но более низкой сахаристостью. Они дают наибольший сбор сахара с гектара, но меньший его выход с единицы массы сырья. Сорта и гибриды сахаристого направления отличаются от первой группы сортов и гибридов меньшей массой корнеплодов, но более высокой сахаристостью, которая у них является более устойчивым признаком и варьирует в пределах 14,4—18,6 % по сравнению с 12,4—17,5 % у сортов и гибридов урожайного направления. Вместе с тем сорта и гибриды урожайно-сахаристого (совмещенного) направления в некоторые годы обеспечивают более высокий сбор сахара с 1 га, чем урожайные или сахаристые.
С 80-х гг. XX в. выращивают только односемянные сорта и гибриды сахарной свеклы. Первый односемянный гибрид урожайно-сахаристого направления Юбилейный — устойчивый к болезням, малоцветушный — был районирован в бывшем СССР в 1981 г.
Среди районированных в настоящее время гибридов преобладают тригоюидные и диплоидные формы. Эти гибриды имеют высокую продуктивность в разных почвенно-климатических условиях, устойчивы к цветушности, корнееду и церкоспорозу.
267
7.1. Анатомо-морфологические особенности растений первого года жизни
Семя. Масса крупных семян сахарной свеклы достигает 5—6 мг, мелких
—около 1,5 мг. При посеве сахарной свеклы используют плодики — односемянные коробочки с сухим и твердым околоплодником, которые при прорастании семени раскрываются путем сбрасывания крышечки. Семена красновато-фиолетовые или темно-вишневые, почковидно-эллипсовидной формы, с заостренным выступом. Семя включает зародыш, центральную его часть — перисперм, содержащий запасные вещества, остаток эндосперма и оболочку.
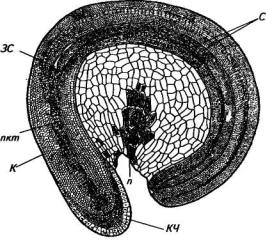
Зародыш представляет собой прикрытое оболочкой кольцевидно согнутое тело, охватывающее перисперм, состоящее из двух сложенных семядолей, связанных через гипокотиль с зародышевым корешком (рис. 81). Перисперм заполнен крахмальными зернами овальной формы.
Гипокотиль покрыт тонкой кутикулой с устьицами, имеет розовую, красную или желтую окраску. На кончике зародышевого корешка находятся верхушечная меристема, прикрытая корневым чехликом, из которой развивается центральный цилиндр корня, первичная кора, кожица и корневой чехлик. В цилиндрической части корешка расположен прокамбий, который тянется по длине
Рис. 81. Зародыш семени свеклы:
п — перисперм; К — корешок, КЧ — корневой чехлик; С — семядоли, пкт — прокамбиальный тяж, ЗС — зачаток стебля
268
корня и переходит в прокамбиальные тяжи двух семядолей, представляющих собой главные проводящие пучки семядолей и будущие сосудистые пучки главного корня.
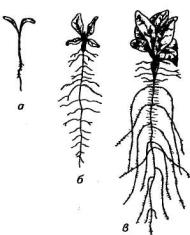
Проросток. Растение в этот период, называемый фазой вилочки, имеет полностью сформированный фотосинтетический аппарат в виде двух содержащих хлоропласты семядолей, главный корень с зачатками боковых корней и проводящие ткани, обеспечивающие транспорт воды и минеральных элементов из почвы в семядоли и ассимилятов из семядолей в корень.
Семядоли продолговато-эллиптические, с тупой верхушкой и вытянутым основанием, переходящим в черешок (рис. 82). Длина семядоли до 2,5 см, ширина 5—7 мм, толщина 0,8—1 мм, ткань ее дифференцирована на палисадную и губчатую паренхиму, а верхняя и нижняя поверхности покрыты эпидермисом со слоем кутикулы, в котором имеются устьица — по 50—60 и 35—45 шт/мм2 соответственно на верхней и нижней сторонах. Через черешок проходит проводящий пучок, связанный с системой жилкования семядоли. Устьица имеются и на гипокотиле, покрытом тонкой кутикулой. Функционируют семядоли до развития 6—8 настоящих листьев, после чего отмирают.
Листья. Первая пара розеточных листьев образуется из зачатков листьев в верхушке стеблевой части меристематического бугорка. Листовые бугорки в последующий период онтогенеза возникают непрерывно, обеспечивая образование новых листьев на протяжении всей вегетации, а при создании благоприятных световых и температурных условий — на протяжении многих лет. Закономерность листообразования у свеклы выражается величиной 5/13; это означает, что через каждые пять оборотов спирали вокруг головки корнеплода на растении образуется 13 листьев, а с 14-го листа начинается новый цикл размещения следующих 13 листьев. На протяжении первого года вегетации свеклы может осуществляться 4—5 таких циклов с образованием 50—60 и более листьев на одном растении.
Форма листовых пластинок первых 6—8 листьев лопатчатая с наибольшей шириной в середине
Рис. 82. Корневая система сахарной свеклы в ранние фазы развития:
а — фаза вилочки; б— утолщение гипокотиля; в — формирование корнеплода
269
пластинки, с тупой верхушкой и клиновидным в направлении черешка основанием. К середине вегетации листья принимают яйцевидную форму с сердцевидным основанием, к концу вегетации они имеют удлиненную лопатчатую форму. Листья средних ярусов обычно гофрированные, что связано с более ранним завершением роста жилок, чем паренхимных тканей в петлях сети жилок.
Площадь листовых пластинок, завершивших рост, также изменяется в онтогенезе: она минимальна у листьев 1—3-го ярусов (в пределах 15—45 см2), достигает максимума в середине вегетации (300—400 см2) и уменьшается к концу вегетации, что определяется изменением напряженности донорноакцепторных отношений и степенью обеспеченности процессов листообразования ассимилятами в растении в течение вегетации. Продолжительность жизни листьев в условиях, близких к оптимальным, наименьшая у первых розеточных листьев (26—37 дней), максимальная у листьев среднего яруса (60 и более дней) и несколько меньше у листьев верхних ярусов. Листовая пластинка сахарной свеклы как мезофитного растения состоит из палисадной и губчатой паренхим, покрытых снаружи эпидермисом и кутикулой. Количество устьиц на верхней стороне листовых пластинок достигает 100 шт/мм2, а на нижней стороне — свыше 150 шт/мм2.
По мере перехода от нижних к верхним ярусам несколько возрастает ксероморфность листьев — уменьшается размер клеток. Поверхность черешка листа также покрыта эпидермисом и кутикулой; две его ткани — колленхима, выполняющая механическую роль, и паренхима, содержащая хлоропласты, тянутся полосами вдоль черешка. Колленхима сильно развита в выступах черешка, заполняя их целиком, а между клетками колленхимы размещаются мелкие паренхимные клетки. Основная же ткань черешка представлена крупноклеточной паренхимой, в которую погружены сосудистые пучки; количество их уменьшается от основания черешка к листовой пластинке. На рост черешков расходуется значительная часть ассимилятов, идущих на рост и развитие листового аппарата: соотношение массы сухого вещества черешков и листовых пластинок варьирует в онтогенезе у разных генотипов в пределах
0,9—1,3.
Корневая система и корнеплод. Они развиваются из зародышевого корешка семени. На ранних фазах роста на центральном корне ниже шейки в плоскости расположения семядолей и первичной ксилемы образуются боковые корешки, усеянные на концах сетью корневых волосков (см. рис. 82). В указанной плоскости по мере роста корня в фазе 2—4 пар листьев первичная кора растрескивается — происходит процесс линьки. Растет корневая система
270