

Кошкин
.pdfну прекращается, когда в генеративной сфере главного надземного побега начинаются процессы микро- и макроспорогенеза. С этого периода линейный рост столона переходит в объемный, т. е. в клубнеобразование.
Клубень — видоизмененный побег, образующийся при разрастании субапикальной части столона. Схематическое изображение тканей на поперечном и продольном срезах клубня Solarium tuberosum показано на рисунке 95. Поверхность клубня покрыта пробкой. Толщина пробки — генетический признак; она варьирует в зависимости от сорта от 50 до 500 мкм, что соответствует 6—19 слоям клеток. В процессе роста клубня суберинизация периферийных клеток перидермы идет от его основания (пуповины) к верхушке.
Одновременно с образованием перидермы под устьицами клубня начинают формироваться чечевички — будущие отверстия в пробковой ткани, обеспечивающие приток кислорода в глубинные ткани. Их количество варьирует от 70 до 150 на один клубень и зависит от размера клубня, типа почвы и погодных условий.
На свежих поперечных или продольных срезах клубня в центре видна толща клеток, образующих сердцевину (см. рис. 95). Периферийную часть сердцевины называют перимедуллярной зоной. Клетки этой зоны интенсивно делятся при разрастании клубня. Отложение в запас ассимилятов в виде крахмальных зе-
Рис. 95. Поперечный (а) и продольный (б) срезы клубня:
1 — пробка; 2 — кора; 3 — проводящие пучки; 4 — сердцевинные лучи; 5 —сердцевина; б— глазок
301
рен осуществляется в паренхимных клетках клубня, которые в совокупности формируют его запасающую ткань. Эта ткань является результатом деятельности перицикла, камбия и клеток перимедуллярной зоны сердцевины. Запасающая ткань составляет почти всю массу клубня. У позднеспелых сортов клетки запасающей ткани крупнее, чем у раннеспелых. Пуповинная часть клубня содержит крахмала больше, чем верхушка, однако по размеру крахмальных зерен пуповинная часть уступает верхушке клубня.
Условия выращивания влияют на формирование тканей клубня, количество и величину крахмальных зерен. Азотные и фосфорные удобрения способствуют увеличению толщины пробки и размера клеток запасающей паренхимы. Нарушение водного режима почвы (засуха или избыток влаги) приводит к уменьшению толщины коры. Хотя объем крахмальных зерен зависит от условий произрастания картофеля, влияние внешних факторов может перекрываться сортовыми признаками. Величина крахмальных зерен коррелирует со зрелостью клубня. Полностью сформированные клубни содержат больше крупных зерен.
Морфофизиологическими характеристиками клубня являются число и расположение глазков. У культурного картофеля преимущественно мелкое заложение глазков, а количество их зависит от сорта. На апикальной части клубня больше глазков, чем на базальной. При разрастании клубня в ширину и толщину увеличивается расстояние между глазками, поэтому шаровидные клубни имеют меньше глазков на единицу массы и единицу поверхности клубня. Клубни с меньшим числом глазков формируют больший урожай, а клубни с большим числом глазков — большее число побегов. В большинстве случаев образуется 3—5 стеблей.
В зависимости от сорта форма клубней сильно различается — от удлиненно-овальной до круглой. Цвет мякоти желтый, кремовый или белый, а кожуры — светлый, фиолетовый, розовый, охристый или красный. Округлые клубни с белой или желтой мякотью, как правило, отличаются большим содержанием сухих веществ в биомассе.
Корневая система картофеля имеет три типа корней: главный, боковой, придаточный. Главный корень развивается при выращивании растений из ботанических семян. В последующем на базальной части стебля сеянца формируются придаточные корни. Если же растение выращивают из клубня, то придаточные корни образуются на нижней части формирующихся стеблей. Первые зачатки корней на ростках клубня появляются, когда росток достигает 3—5 мм толщины. Диаметр корней картофеля от 1 до 3 мм. При проращивании клубней на свету корни начинают ветвиться
302
на ранней стадии формирования, и при посадке в почву образуется мощная корневая система.
На столонах по мере их роста в узлах формируются столонные (придаточные) корни диаметром 0,3—1 мм. Количество столонных корней зависит от влажности почвы, при благоприятном увлажнении их число увеличивается.
Основная масса корней (60—80 %) находится в пахотном слое. Позднеспелые сорта образуют более мощную и глубже проникающую корневую систему, чем раннеспелые. Между массой корней, глубиной их проникновения в почву и массой ботвы существует положительная корреляция.
Цветки собраны в соцветие, представляющее собой сложный завиток, расположенный на общем цветоложе различной длины. Склонность к цветению зависит от сорта и фотопериодических условий.
Семя состоит из зародыша, эндосперма и оболочки. Зародыш состоит из двух семядолей, гипокотиля и корешка; окружен эндоспермом, клетки которого содержат жиры и белки.
Плод — двугнездная ягода, шаровидная, овальная или коническая, образована двумя плодолистиками. Из ткани плаценты образуется мякоть зрелого плода, в которую погружены семена.
Картофель сравнительно редко выращивают из ботанических семян, в основном в селекционно-генетических целях. При прорастании семени семядоли выносятся на поверхность почвы за счет удлинения гипокотиля. Зародышевый корень развивается как главный, который вскоре образует боковые корни. Первые настоящие листья овальные, покрыты волосками. Когда растение достигает высоты всего нескольких сантиметров, в пазухах семядолей начинают формироваться столоны. Столоны врастают в почву и образуют мелкие клубни.
До появления всходов и создания органов автотрофного питания материнский клубень служит источником энергопластических веществ для ростовых процессов. Прорастание начинается с верхних глазков, причем в рост трогается обычно одна почка глазка. Если появившиеся ростки обломать, то в этих же глазках пробуждается вторая почка, а если удалить и эту, то прорастает следующая. На образование ростков затрачиваются пластические вещества. Обламывание ростков проросших клубней при посадке отрицательно сказывается на росте и развитии картофеля.
Поврежденные клубни прорастают быстрее, чем неповрежденные, из-за потери вместе с кожурой ингибиторов роста, более интенсивного дыхания. Щадящая уборка и сортировка, низкие температуры при хранении (2—3 °С) способствуют поддержанию состояния покоя клубня. С помощью различных химических веществ можно регулировать длительность периода покоя клуб-
303
ней, влиять на физиологическое состояние посадочного материала. Органогенез картофельного растения включает 12 этапов.
Этап |
Характеристика развития растении |
I |
Прорастание клубня или семени, при котором конус нарастания |
зачаточного стебля имеет форму гладкого овального бугорка |
|
II |
Заложение почек, формирующих подземные побеги |
III |
Формирование бугорков зачаточных узлов и междоузлий будущего |
|
соцветия |
IV |
Образование на зачаточной оси соцветия цветкового бугорка |
V |
Начало образования цветка, формирование в пыльниках |
археспориальной ткани |
|
VI |
Начало репродукции, процессы микро- и макроспорогенеза; индукция |
процесса разрастания клеток тканей субапикальной части столона |
|
VII |
Формирование одноядерной пыльцы (гаметогенез); рост покровных |
частей цветка |
|
VIII |
Формирование двуядерной пыльцы; формирование бутона (бутонизация) |
IX |
Оплодотворение и образование зиготы (цветение) |
X |
Определяется строение ягоды и семени, характерное для картофеля |
XI |
Продолжение процесса дифференцировки зародыша и эндосперма, |
накопление питательных веществ в семени |
|
XII |
Завершение дифференцировки зародыша и эндосперма |
Почки, формирующие подземные побеги, закладываются на II этапе органогенеза, когда конус нарастания монокарпического ортотропного побега имеет 2—5 зачатков листьев на 2—5 узлах метамеров. Согласно Ф. М. Куперман, на этом этапе онтогенеза конус нарастания определяет вегетативную сферу (ветвление) растения. Почки боковых побегов надземной части стебля формируются после почек специализированных побегов вегетативной репродукции.
Как генетически детерминированный органообразующий процесс, клубнеобразование проявляется в оптимальных для роста и развития условиях на определенном этапе органогенеза. Объемный (радиальный) рост в субапикальной части столона начинается на VI этапе органогенеза, когда в генеративной сфере главного ортотропного побега идут процессы микро- и макроспорогенеза. С переходом к объемному росту линейный рост столона прекращается.
Следует отметить, что не все столоны формируют клубни. Клубнеобразующие столоны отличаются лучшим развитием ксилемы, большим числом и размером крахмальных зерен (табл. 63). Это свидетельствует о более интенсивном притоке ассимилятов в субапикальную часть клубнеобразующих столонов.
304
63. Характеристика столонов картофеля сорта Приекульский ранний
Показатель |
|
Столоны |
|
клубнеобразующие |
|
неклубнеобразующие |
|
|
|
||
Количество сосудов ксилемы, шт/мм |
1683 ±87 |
|
827 + 69 |
Толщина вторичной ксилемы, мкм |
111,6 ± 2,8 |
|
Вторичная ксилема не развита |
Диаметр крупного сосуда, мкм |
49,3 ±3,5 |
|
18,5 ±2,8 |
Число крахмальных зерен в одной |
35,0 + 2,9 |
|
6,5 ±0,4 |
паренхимной клетке флоэмы |
|
||
|
|
|
|
Величина крахмальных зерен в |
26,0+1,7 * |
|
2,4 ±0,4 |
паренхимных клетках флоэмы, мкм |
|
||
|
|
|
|
Подавляющее большинство селекционных сортов вида картофеля S. tuberosum проявляют нейтральную фотопериодическую реакцию клубнеобразования. Это означает, что процесс перехода столона в клубень не зависит от продолжительности дня. Лишь у небольшой группы сортов под воздействием короткого фотопериода накопление массы клубней усиливается.
Клубнеобразование — сложный процесс, включающий образование столонов, индукцию и образование клубней, их дальнейший рост и созревание. Как уже отмечалось, почки, формирующие подземные побеги (столоны), закладываются на II этапе органогенеза на базальных узлах ортотропного побега. Начало заложения почек, формирующих столоны, не зависит от фотопериода.
Переходу столона в клубень предшествует дедифференцировка клеток постоянных тканей в субапикальной части столона. У короткодневных по клубнеобразованию видов картофеля этот процесс происходит только в условиях короткого дня при воздействии на клетки фактора, образующегося в листьях. Природа этого фактора до сих пор не установлена. У нейтральной группы видов, форм и сортов картофеля процесс образования клубней детерминирован, так как образование листового фактора не зависит от продолжительности фотопериода. По-видимому, у этих растений реализуется принцип автономного механизма, аналогичный установленному М. X. Чайлахяном для процесса цветения. У промежуточных (количественных по фотопериодической реакции клубнеобразования) растений образование листового фактора, по-видимому, происходит и в условиях длинного дня, но с низкой интенсивностью света. Короткий фотопериод лишь усиливает этот процесс.
Гормоны влияют на состояние субапикальной зоны столона, которая, разрастаясь радиально, переходит в клубень. Начало разрастания столонов характеризуется повышением уровня активности цитокининов и АБК на фоне достаточно высокой активности ИУК. Известные фотогормоны, по-видимому, не являются непосредственными индукторами клубнеобразования, но регулируют
305
деление и рост клеток, дифференциацию тканей, способствуют созданию аттрагирующей силы, являющейся основой притока ассимилятов и воды в клубни.
8.2. Формирование и функциональная активность фотосинтетического аппарата растений
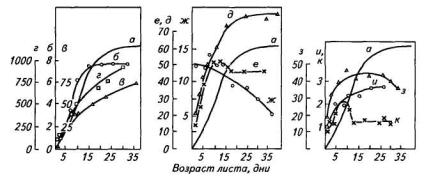
Формирование мезоструктуры фотосинтетического аппарата и развитие функциональной активности листьев картофеля подробно исследовано А. Т. Мокроносовым с сотрудниками. В качестве модели использовали верхушечную долю листа среднего яруса (7-й лист, считая от основания побега) от начала развертывания до отмирания. Продолжительность роста этого листа у картофеля сорта Малахит составляла 26—28 дней, и после этого он оставался функционально активным еще 10—12 дней (рис.96). Первые 10—12 дней рост листа происходит за счет деления и роста клеток, а затем только путем растяжения клеток без увеличения их числа. Число хлоропластов в клетке увеличивается до тех пор, пока возрастает объем клетки. В онтогенезе листа относительный объем всех хлоропластов объема клетки постепенно снижается от 25—30 до 3—5 % объема клетки. Максимальный объем одного хлоропласта составляет 40—50 мкм3. Объемы хлоропластов достаточно стабильны и мало подвержены фенотипическим изменениям.
Рис. 96. Динамика структурных характеристик листа картофеля сорта Малахит в онтогенезе (по А. Т. Мокроносову):
а —площадь листа (см2); б— общее число клеток (106) в листе; в —объем клетки палисадной ткани (тыс. мкм3); г —объем клетки, соответствующей одному хлоропласту (мкм3); д — число хлоропластов в клетке палисадной ткани; е — число хлоропластов в клетке губчатой ткани; ж — число хлоропластов (106/cм2), з — объем хлоропласта (мкм3); и — содержание хлорофилла (109 молекул/хлоропласт); к — индекс проективной поверхности хлоропластов
306
Исключение составляет действие затенения, которое индуцирует значительное (в 2 раза) увеличение объема пластид. Под влиянием дефицита света увеличивается количество антенного хлорофилла, обеспечивающего реакционный центр.
По мере роста листа число пластид на 1 см2 площади снизилось с 14 до 6 млн. Фенотипическое варьирование этого показателя у картофеля не превышало 1,3—1,7-кратности. Например, при оптимальном водоснабжении и сильной засухе количество пластид составляло соответственно 15,6 и 25 млн/см2. Дефицит воды приводит к мелкоклеточности из-за задержки растяжения клеток. При этом число пластид в клетке снижается меньше, чем объем клеток.
Синтез хлорофилла в листе продолжается и после достижения хлоропластами своего максимального объема. К завершению роста листа картофеля содержание хлорофилла в одном хлоропласте составляет в среднем 0,5 • 109 молекул. Хлоропласты палисадной ткани содержат меньше хлорофилла, но фотохимическая активность этого пигмента выше, чем в хлоропластах губчатой ткани.
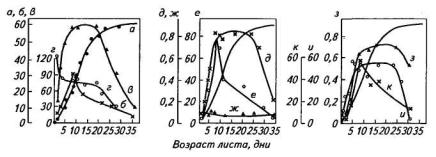
Суммарная поверхность наружных мембран хлоропластов достигает наибольшей величины при площади листа 60 % от конечной и составляет 10 дм2/дм2 листа. При расчете на единицу листовой поверхности наибольшая интенсивность фотосинтеза наблюдается в листьях, достигших 0,4—0,6 от максимальной площади (рис. 97). Следовательно, максимальному фотосинтезу соответствует во времени и наибольшая величина общей и проективной поверхностей хлоропластов.
Рис. 97. Динамика функциональных характеристик листа картофеля сорта Малахит в онтогенезе:
а —площадь листа (см2); б— фотосинтез мг СО2/(дм2ч); в —фотосинтез единичного хлоропласта [106 молекул/(хлоропласт∙сек)]; г —фотосинтез (моль СО2/(моль хлорофилла∙ч); д — активность РБФ-карбоксилазы (мкмоль СО2/(мг белка • мин); е — то же (мкмоль СО2/ (дм2 • мин); ж — активность ФЕП-карбоксилазы (мкмоль субстратаДмг белка∙мин); з — активность НАДФ-ГАФ-дегидрогеназы [мкмоль субстратаДмг белка∙мин)]; и —активность РБФ-карбоксилазы [мкмоль CO2/109 (хлоропластов∙мин)]; к — активность НАДФ-ГАФ- дегидрогеназы (мкмоль субстрата /(дм2∙мин)
307
Расчеты фотосинтеза на хлорофилл — ассимиляционное число (АЧ) — показывают, что за время жизни листа величина АЧ снижается в 4—5 раз: с 15 до 3 мг СО2/(мг хлорофилла • ч). Это связано с накоплением по мере старения листа фотохимически неактивных форм хлорофилла и увеличением количества антенного хлорофилла в фотосинтетической единице.
Активность ключевого фермента фотосинтеза — РБФ-карбоксилазы — быстро нарастает в ювенильном листе и сохраняется высокой вплоть до завершения роста листа. С возрастом общее содержание белка и ферментных белков снижается, поэтому при пересчете на площадь листа наибольшая активность ферментов, как и интенсивность фотосинтеза, обнаруживается в листе с площадью 40—50 % от максимальной.
Структурная организация, активность ферментов, фотохимическое участие хлорофилла и ряд других физиологических характеристик коррелируют с направленностью углеродного метаболизма в онтогенезе листа. На ювенильных фазах доминирует карбоксилирование ФЕП. На экспоненциальном участке роста листа происходит интенсивное новообразование РБФ-карбоксилазы. В зрелом листе СО? фиксируется почти полностью на РБФ, активность ФЕП-карбоксилазы составляет лишь около 10 % от максимальной активности РБФ-карбоксилазы. Возрастные особенности фотосинтетического метаболизма соответствуют характеру дальнейшего превращения ассимилятов: ювенильный лист получает амино- и оксикислоты, используемые в биосинтезе растущего листа, а зрелый лист — сахарозу, т. е. основную транспортную форму углеводов.
По типу фотосинтетического метаболизма картофель — классический С3- вид. Первичным продуктом фиксации СО2 является ФГК. До 95 % поглощаемого при фотосинтезе СО2 используется на синтез сахарозы и крахмала. Листья картофеля имеют сравнительно низкую величину фотодыхания — около 20 % от фотосинтеза. Углекислотный компенсационный пункт составляет около 0,004 %.
8.3.Водный режим
Вотличие от многих других культур хозяйственно полезная часть урожая картофеля формируется под поверхностью почвы. При рассмотрении вопроса о водообеспеченности картофеля и оптимальной влажности почвы следует учитывать высокую чувствительность молодых клубней к аэрации, которая зависит от типа почвы и снижается по мере повышения содержания в ней влаги. Снижение концентрации О2 в среде до 10—15 % приводит к резкому угнетению роста клубней. Оптимальное содержание
308
влаги для роста растений и образования высокого урожая клубней на легких почвах — 75—80 % НВ, средних — 70 и тяжелых — 50-60% НВ.
Осадки в первой половине вегетации усиливают рост ботвы, в период бутонизации положительно влияют на число клубней в гнезде, во второй половине вегетации — на массу клубней. Высокие урожаи картофеля получают на культурных почвах при 300 мм осадков, большая часть которых выпадает в период бутонизации и начала формирования клубней и обеспечивает влажность почвы после бутонизации, равную 70—75 % НВ. Дефицит влаги в период от начала клубнеобразования до созревания клубней может существенно тормозить процесс формирования урожая. Имеются сведения, что при дефиците влажности почвы в 1 см недобор урожая клубней может достигать 3 т/га. При снижении водного потенциала с —0,5 до —1,5 • 102кПа урожайность уменьшалась с 28 до 20 т/га. В таких условиях орошение в начале клубнеобразования положительно сказывается на приростах массы клубней и ускоряет сроки уборки урожая. На легких песчаных почвах еженедельный полив полной нормой давал прибавку урожая до 1,5 т/га на каждый сантиметр воды.
Запасы влаги в материнском клубне, которые увеличиваются после высадки, обеспечивают появление всходов и проникновение корневой системы в более увлажненные слои почвы. С развитием корней запасы воды материнского клубня играют роль страхового фонда, способного возместить недостаток почвенной влаги в наиболее напряженные часы суток. В дальнейшем эту роль могут выполнять и молодые клубни, оводненность которых в течение суток при засухе значительно колеблется. Уменьшение оводненности молодых клубней в дневные часы обусловлено мобилизацией воды в побеги, особенно в листья. Поэтому даже в засушливые периоды часто не наблюдают значительного подвядания ботвы картофеля, тогда как листья сахарной свеклы теряют тургор.
Содержание воды в листьях картофеля зависит от водоснабжения и условий, определяющих интенсивность транспирации. Нижние листья более оводнены, чем верхние, но по мере старения растений количество воды в них также снижается. Дефицит насыщения тканей водой отмечается чаще в полуденные часы и может достигать 20—25 %. Важный показатель водообмена
— интенсивность транспирации — зависит от напряженности метеорологических факторов и контролируется устьичным аппаратом. Суточный ход скорости транспирации листьев картофеля в районах с достаточным влагообеспечением и умеренными температурами характеризуется одновершинной кривой с максимумом в полуденные часы. В районах с ограниченным водоснабжением часто наблюдается двувершинная кривая. Скорость транспирации в полу-
309

денные часы снижается вследствие частичного закрывания устьиц, что способствует сокращению потерь влаги и сохранению оводненности листьев. Содержание воды в листьях картофеля сравнительно высокое — 80—90 %.
В южных районах наибольшие величины транспирации [190—290г/(м2ч)] отмечены в фазе бутонизации — цветения, в северных —до бутонизации [300— 360 г/(м2∙ч)]. Северные растения отличаются более высокой скоростью транспирации, оводненностью тканей, незначительными колебаниями оводненности в течение суток и вегетации.
Расход воды на испарение определяется габитусом растения, величиной его листовой поверхности. Транспирационный коэффициент (ТК) — расход воды на создание единицы сухой массы — может колебаться в широких пределах в зависимости от сорта и места произрастания. Среди других культур картофель занимает среднее положение по величине транспирационного коэффициента (рис. 98). Величины ТК, характерные для северных условий, выше, чем в южных районах, а продуктивность транспирации в 2—3 раза ниже. За вегетацию потребление воды высокопродуктивными ценозами картофеля в средней зоне составляет 2000— 2800 т/га. В южных районах и Поволжье, где осадков выпадает меньше, картофель весьма отзывчив на орошение.
Важно отметить, что температурный оптимум клубнеобразования повышается с увеличением влагообеспеченности почвы. Установлено, что на подзолистых суглинках при запасах продуктивной влаги в пахотном горизонте около 50 мм температурный оптимум находится в пределах 14—18 °С, а при 20 мм — снижается до 11—12 °С. Следовательно, регулируя запасы влаги, можно
обеспечить |
близкие |
к |
оптимальным |
|
температурные |
условия |
||||
клубнеобразования. |
|
|
|
|
|
|
|
|
|
|
Избыточное увлажнение почвы, как и недостаток влаги, сильно влияет на |
||||||||||
|
|
|
|
водный |
обмен |
и |
продуктивность |
|||
|
|
|
|
картофеля. Клубни получаются мелкими, |
||||||
|
|
|
|
с низким содержанием сухого вещества и |
||||||
|
|
|
|
крахмала, |
|
усиливается |
|
поражение |
||
|
|
|
|
растений болезнями. Нарушения водного |
||||||
|
|
|
|
режима приводят к торможению роста |
||||||
Рис. 98. Средние величины |
|
клубней. При переувлажнении почвы |
||||||||
|
наблюдается |
|
резкое |
|
|
снижение |
||||
транспирационного коэффициента |
транспирации, |
|
|
уменьшается |
||||||
различных культур: |
|
|
оводненность |
тканей, |
|
увеличивается |
||||
1 — сахарная свекла, кукуруза; 2— |
|
|
||||||||
|
водный дефицит листьев, изменяется |
|||||||||
пшеница, ячмень; 3— картофель, |
|
|||||||||
подсолнечник; 4— рапс, горох; 5 — |
|
соотношение |
свободной |
|
и |
связанной |
||||
|
люцерна |
|
|
воды (табл. 64). |
|
|
|
|
||
310