

Кошкин
.pdf64. Влияние переувлажнения и почвенной засухи на водный режим листьев картофеля
|
|
Интенсивность |
Содержание |
Отношение |
|
|
|
Влажность |
свободной |
Водный |
|||
Вариант опыта |
транспирации, |
воды в |
||||
почвы, % НВ |
воды к |
дефицит, % |
||||
|
2 |
|
||||
|
|
г/(м ∙ч) |
листьях, % |
связанной |
|
|
Контроль |
65-70 |
320 |
82 |
0,21 |
8 |
|
Затопление, сут: |
|
|
|
|
|
|
3 |
115 |
65 |
77 |
0,10 |
10 |
|
7 |
109 |
70 |
72 |
0,11 |
24 |
|
Засуха, сут: |
|
|
|
|
|
|
3 |
60 |
70 |
80 |
0,10 |
11 |
|
7 |
50 |
65 |
79 |
0,07 |
17 |
Избыточное увлажнение вызывает нарушение общего метаболизма, усиливает анаэробное дыхание корневой системы. В результате снижается интенсивность поглощения основных элементов минерального питания, в частности азота. Это тормозит синтез и обновление зеленых пигментов, белков, приводит к нарушению хлорофилл-белковых комплексов. Существенно снижаются продуктивность работы единицы хлорофилла и прирост биомассы растений (табл. 65).
65. Влияние влажности почвы на продукционный процесс картофеля (на восьмой день опыта)
Влажность |
Содержание |
Среднесуточная |
Сухая масса, г/растение |
|||
продуктивность |
|
|
|
|
||
почвы, % |
хлорофилла, |
хлорофилла, мг |
общая |
листьев |
стеблей |
клубней |
НВ |
мг/растение |
сухой массы/мг |
||||
|
|
хлорофилла |
|
|
|
|
63 |
122 |
34 |
39,1 |
12,7 |
5,5 |
18,3 |
109 |
58 |
3 |
16,4 |
8,0 |
7,1 |
— |
Таким образом, воздействуя на процессы жизнедеятельности и активность фотосинтетического аппарата, водный фактор играет важную роль в формировании продуктивности картофеля. На урожайности неблагоприятно сказываются как недостаточное, так и избыточное увлажнение почвы.
8.4. Фотосинтез
При изучении фотосинтеза картофеля особое внимание уделяли поиску генотипов, отличающихся высокой фотосинтетической активностью.
311
Анализ структуры и функциональной активности листьев различных видов и сортов картофеля показал, что по общей площади листьев сорта в десятки раз отличались от диких и культурных видов. Однако по большинству структурно-функциональных показателей достоверных различий не выявлено. Активность хлоропластов выше у селекционных сортов, но не настолько, чтобы эти различия могли сильно повлиять на продуктивность. Высокая продуктивность селекционных сортов обусловлена прежде всего отбором форм с более мощным листовым аппаратом (площадь листовой поверхности).
Вместе с тем некоторым исследователям среди множества клонов картофеля удалось выявить генотипы, отличающиеся повышенной (более чем в 1,5 раза) фотосинтетической активностью. Эти клоны способны поддерживать повышенную интенсивность фотосинтеза в течение дня и вегетационного периода и характеризуются высокой продуктивностью.
О варьировании интенсивности фотосинтеза свидетельствуют данные таблицы 66. Различия в интенсивности видимого фотосинтеза между видами и сортами в расчете на единицу площади листьев выражены довольно сильно. Однако при пересчете фотосинтеза на единицу сухой массы различия между ними существенно сглаживались. Это связано с разной удельной поверхностной плотностью листьев. Листовые пластинки селекционных сортов крупнее и тоньше двух других видов, а их общая площадь существенно выше.
66. Интенсивность фотосинтеза листьев среднего яруса растений разных видов и сортов картофеля при температуре 20—22 °С (июль)
Вид, сорт |
Фотосинтез |
|
мг СО2 /(дм2 • ч) |
мг СО2 /(г • ч) |
|
S. tuberosum: |
|
|
Идеал |
26,7 ±1,0 |
51,1 ±1,9 |
Искра |
16,1 ±2,6 |
44,6 ±8,9 |
Шарташский |
12,9 ±1,4 |
55,4 ±5,0 |
Приекульский ранний |
- |
56,0 ±4,3 |
S. andigenum |
39,2 ±2,8 |
58,0 ±4,2 |
S. stoloniferum |
37,8 ±1,7 |
49,6 ±2,2 |
Характер возрастных кривых ИФ листьев разного яруса иллюстрирует рисунок 99. Наибольшие величины ИФ — до 60 мг СО2/(г • ч) были зарегистрированы у листьев средних ярусов. Верхушечные листья фотосинтезировали слабее.
312

Рис. 99. Возрастные изменения фотосинтетической активности листьев картофеля сорта Приекульский ранний. (Цифрами около кривых обозначен номер листа, считая от основания побега)
Для полной реализации фотосинтетической способности растений условия среды должны быть оптимальными. Листья картофеля насыщаются при освещенности 60—80 % от полной солнечной. В умеренной зоне для фотосинтеза оптимальна температура 15—25 °С. Растения сорта Русеет Фурбанк, выращиваемые при орошении в условиях пустыни на юге Айдахо (США), при варьировании суточной температуры от 16 до 35 °С имели температурный оптимум фотосинтеза в пределах 24—28 °С. Адаптированный к северным условиям сорт картофеля Приекульский ранний способен фотосинтезировать со средней интенсивностью 10— 15 мг СО2/(г • ч) при 10— 20 °С, обеспечивая за день ассимиляцию до 150—260 мг СО2/дм2 листьев.
Дневная динамика ИФ листьев картофеля при достаточном содержании почвенной влаги в ясные дни имеет форму одновершинной кривой либо следует за ходом освещенности. В южных районах при повышенных температурах и недостатке влаги возможна дневная депрессия фотосинтеза, соответствующая наиболее высокой температуре дня.
Что касается фотосинтетических пигментов, то максимум содержания хлорофиллов (1,7—2 мг/г сырой массы) и каротиноидов (0,4—0,6 мг/г) отмечен в фазе бутонизации. Самыми высокими концентрациями хлорофиллов отличаются листья среднего и верхнего ярусов. В период налива клубней листья содержат в 2—3 раза меньше пигментов, что свидетельствует о снижении скорости их обновления. Соотношение хлорофиллов а и ft до фазы налива клубней сравнительно высокое и варьирует в пределах 3,3—4,5, затем заметно снижается. Доля хлорофиллов светособирающего комплекса (ССК) составляет 40—45 %.
Содержание хлорофиллов в листьях картофеля сортоспецифично (табл. 67). Ранние и среднеранние сорта отличаются более низким содержанием пигментов по сравнению со средне-поздними и поздними. Минимальное содержание хлорофиллов отмечено у примитивных культурных видов. У разных видов картофеля количество хлорофилла в хлоропласте колеблется от 0,3—0,5 • 109 до 5 • 109 молекул/хлоропласт. Установлена тесная связь между содержанием хлорофиллов в листьях и крахмалистоcтью клубней.
313
67. Содержание пигментов и доля хлорофиллов светособираюшего комплекса (ССК) в листьях различных сортов и примитивных видов картофеля в фазе бутонизации, мг/г сухой массы
Сорт |
|
|
Хлорофилл |
|
ССК, % |
Сумма |
||
|
а |
|
в |
|
а/в |
каротинои- |
||
|
|
|
|
|
дов |
|||
|
|
|
|
|
|
|
|
|
|
|
Ранние сорта |
|
|
|
|||
Приекульский ранний |
|
1,4 |
|
0,21 |
|
5,4 |
34 |
0,44 |
Искра |
|
1,04 |
|
0,27 |
|
3,8 |
45 |
0,43 |
|
|
Среднеранние сорта |
|
|
|
|||
Шарташский |
|
0,98 |
|
0,28 |
|
3,5 |
49 |
0,35 |
Сахалинский |
|
1,26 |
|
0,31 |
|
4,1 |
43 |
0,52 |
|
|
Среднепоздние сорта |
|
|
|
|||
Берлихинген |
|
1,54 |
|
0,32 |
|
4,8 |
38 |
0,49 |
Идеал |
|
1,66 |
|
0,36 |
|
4,6 |
39 |
0,58 |
|
Примитивные культурные виды |
|
|
|||||
Pg 1200 |
|
0,95 |
|
0,22 |
|
4,3 |
41 |
0,32 |
Pg637 |
|
0,87 |
|
0,23 |
|
3,8 |
46 |
0,36 |
Хлорофилльный индекс (произведение содержания хлорофилла в фотосинтезирующих органах растения на их сухую массу в расчете на 1 га) посевов картофеля может достигать 20 кг/га, но в среднем варьирует от 12 до 18 кг/га. Для большинства культурных фитоценозов, особенно злаковых, величина ХИ может быть существенно выше. Однако в отличие от них посевы картофеля отличаются сравнительно высокими значениями продуктивности хлорофилла, превышающими 200 кг углерода/кг хлорофилла.
Картофель занимает промежуточное положение среди других культур по средней за вегетационный период величине чистой продуктивности фотосинтеза, которая составляет 3—8 г/(м2∙сут). Однако для него зарегистрированы и максимально высокие значения ЧПФ, равные 15—25 и даже 60 г/(м2∙сут).
Для фотосинтетической продуктивности существенны размеры листовой поверхности, которые определяют поглощение радиации посевом. ИЛП в посевах картофеля может достигать 6—7 м2/м2. Оптимальными в Нечерноземной зоне считаются значения ИЛП 4— 6 м2/м2. В более северных районах для сорта Приекульский ранний оптимум ИЛП равен 3—4м2/м2. Зависимость урожая от площади листьев прямолинейная, если последняя не превышает оптимальную величину.
Для обеспечения формирования урожая клубней ассимилятами большое значение имеет продолжительность фотосинтетической деятельности посева в течение вегетационного периода или фотосинтетический потенциал [ФП, млн (м2∙сут)/га]. На
314
1000 единиц ФП в высокопродуктивных посевах картофеля формируется 20—22 кг клубней. На основании обобщения литературных данных X. Г. Тооминг предложил уравнение регрессии, связывающее величину ФП с урожаем (У, т/га):
У=16,5ФП + 4,3.
Снижение величины ФП в результате повреждения листовой поверхности болезнями или заморозками — одна из причин недобора урожая.
Для многих культур направленность продукционного процесса на формирование урожая удобно оценивать по величине показателя Кхоз. Установить величину Кхоз для картофеля трудно, так как для полной реализации потенциала продуктивности требуется, чтобы все временно депонированные ассимиляты из ботвы транспортировались в клубни, а также максимально полно реутилизировались в молодые клубни продукты гидролиза стареющих надземных органов. Поэтому величину Кхоз целесообразно определять как отношение массы клубней при уборке к максимальной массе ботвы. Величина Кхоз колеблется в широком диапазоне, но основная часть данных находится в пределах 0,8—1,2. Это свидетельствует о высокой эффективности использования продуктов фотосинтеза на формирование урожая у данной культуры.
8.5. Дыхание
Ассимилированный в процессе фотосинтеза углерод включается в биомассу растения при участии дыхания. Ежесуточно в процессе дыхания растения окисляют значительное (30—70 %) количество ассимилированного при фотосинтезе углерода. В дыхании образуются промежуточные продукты (метаболиты), вовлекаемые в общий обмен, и происходит передача энергии на осуществление биосинтезов. Согласно современным представлениям, дыхание обеспечивает энергетически трансформацию субстрата (ассимилятов) в биомассу и поддержание функциональной целостности биомассы. Поэтому изучение дыхания и его роли в продукционном процессе актуально с позиции не только теории, но и практики.
Для растений картофеля характерен выраженный градиент дыхательной активности. Каждый последующий лист (считая от основания побега к верхушке) характеризуется более высокой интенсивностью дыхания по сравнению с расположенным ниже. Наиболее интенсивно дышат самые молодые (верхушечные) листья (рис. 100). Максимальные величины ИД, составляющие 6—8мг/(г сухой массы • ч) (при 20 °С), были отмечены в период, когда листья имели еще низкую интенсивность фотосинтеза и росли преиму-
315
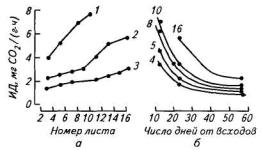
Рис. 100. Дыхательная активность листьев картофеля сорта Приекульский ранний в зависимости от их положения на стебле (а) и возраста (б):
1—3— фаза вегетативного роста, цветения и налива клубней соответственно; 4—16 — номер листа, считая от основания побега
щественно за счет импорта ассимилятов. Когда листья сформировали 40 % площади и стали нетто-экспортерами ассимилятов, их дыхание
снизилось в 2—3 раза. Скорость выделения СО2 у завершивших рост листьев составляет в среднем 1,5—2 мг СО2/(г сухой массы • ч) (табл. 68). По сравнению с другими органами картофеля листья отличаются наибольшей дыхательной активностью.
68. Интенсивность дыхания органов картофеля при температуре 18—22 °С
Орган растения |
ИД, мг СО2/(г сухой массы∙ч) |
Литературный источник |
|
Листья: |
|
|
|
молодые |
7-11 |
Winkler, 1971 |
|
6-9 |
Шевелуха, Маханько, 1971 |
||
|
8-11 |
Головко, 1984 |
|
зрелые |
3-5 |
Winkler, 1971 |
|
3,3 |
Шевелуха, 1980 |
||
|
2-3 |
Головко, 1984 |
|
Листья + стебли |
1,7-2,9 |
Sale, 1974 |
|
Стебли: |
|
|
|
молодые |
1,8 |
Winkler, 1971 |
|
3,5-4,5 |
Головко, 1984 |
||
|
|||
зрелые |
0,5 |
Winkler, 1971 |
|
1,0 |
Головко, 1984 |
||
|
|||
Корни, зрелые |
13-20* |
Bushnell, 1956 |
|
0,6 |
Шатилов, Назарян, 1978 |
||
|
1-3 |
Головко, 1984 |
|
Корни + клубни |
0,24-11,1 |
Sale, 1974 |
|
Клубни: |
0,4-0,8 |
Winkler, 1971 |
|
зрелые |
0,03-0,4 |
Шатилов, Назарян, 1978 |
|
0,7-1,6 |
Головко, 1984 |
||
|
0,6-0,7 |
Головко, 1986 |
|
молодые |
0,9-1,6 |
То же |
*Переведена из мл О2.
316
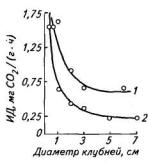
Дыхание стеблей и корней составляет в среднем 1—3 мг СО2/(г∙ч), а клубней — на порядок ниже, чем листьев. Скорость выделения СО2 в зачатках клубней (диаметр менее 1 см) составляет около 2 мг/(г сухой массы∙ч) (при 20 °С). По мере роста скорость дыхания снижается до более или менее постоянного уровня (рис. 101). Клубни диаметром 3—7 см дышат с интенсивностью 0,6—0,7 мг СО2/(г сухой массы∙ч). Такой ход дыхания клубней отражает в первую очередь процессы, связанные с ростом. Когда рост столона в длину прекращается и начинается развитие клубня, главную роль в росте клубня играет увеличение размера клеток. Увеличение размера клеток заметно уже у клубней диаметром 1 см. При достижении клубнями сырой массы 30—40 г, что соответствует диаметру около 3 см, этот процесс становится основным. Таким образом, дыхательная способность клубней, как и других органов, снижается по мере их роста.
Рост клубней осуществляется за счет продуктов текущего фотосинтеза, поступающих из листьев и ранее депонированных в стеблях или других частях растения. Если с помощью полной дефолиации растения лишить формирующиеся клубни (диаметр 1,5—2 см) субстрата, то их дыхание на 3—4- е сут после удаления надземной части стабилизируется на более низком уровне и составит около 0,3 мг СО2/г сухой массы • ч (см. рис. 101). Эта величина сопоставима с интенсивностью выделения СО2 у зрелых клубней в состоянии покоя при хранении.
Картофель занимает ведущее место в группе растений со сравнительно высокой активностью дыхательного метаболизма листьев. Сравнительно высокая интенсивность дыхания листьев в сочетании с достаточно высокой облиственностью приводит к тому, что в течение значительной части вегетационного периода в дыхании целого растения картофеля доминируют листья (рис. 102). Вклад клубней в выделение СО2, несмотря на быстрое нарастание массы, сравнительно невелик, что связано с их низкой дыхательной активностью. Причина столь существенной (на порядок) разницы в скорости дыхания листьев и клубней обусловлена общим уровнем и направленностью метаболизма. Зрелые функционально активные листья содержат в среднем около 40 мг белкового азота, что эквивалентно концентрации белка, равной 250 мг/г сухой массы. Белки листьев представлены преимущественно (80%) растворимыми фракциями, что свидетельствует об их ферментной природе и высокой скорости обновления.
Содержание белка в
Рис. 101. Интенсивность дыхания клубней интактных (7) и дефолиированных (2) растений картофеля сорта Приекульский ранний
317
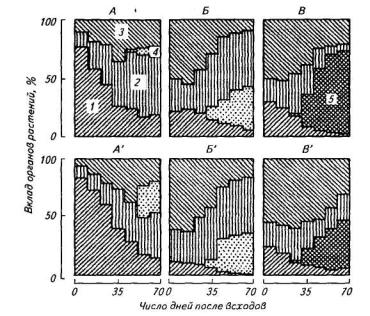
Рис. 102. Доля органов в сухой биомассе целых растений Trifolium pratense (A), Avena saliva (Б) и Solatium tuberosum (В). А'— В'— вклад органов в дыхание целых растений:
1—корни; 2— стебли; 3 — листья; 4— генеративные органы; 5— клубни
клубнях не превышает 35 мг/г сухой массы, что почти на порядок ниже, чем в листьях. Основную же часть сухой массы клубней составляют углеводы (крахмал).
|
ИД листьев средних ярусов растений разных видов |
Вид, сорт |
и сортов картофеля в фазе бутонизация — цветение, |
S. tuberosum: |
мг COj/fe сухой массы • ч) при 20 °С |
|
|
Идеал |
4,5 + 0,2 |
Искра |
3,7 ±0,4 |
Шарташский |
4,6 ±0,5 |
Приекульский ранний |
3,5 ±0,3 |
S. andigenum |
4,5 ±0,2 |
S. stoloniferum |
4,1 ±0,2 |
Поскольку затраты на синтез и обновление белков — одна из наиболее существенных статей расхода энергии и промежуточных метаболитов, поставляемых дыханием, не удивительно, что листья
318
и клубни сильно различаются по дыхательной способности и величине коэффициентов дыхания (табл. 69). Различия в химическом составе биомассы и показателях дыхания обусловливают разную эффективность роста листьев и клубней. Данные, представленные в таблице 70, показывают, что существуют значительные различия между надземными и специализированными органами запасания по величине показателя, характеризующего эффективность превращения ассимилятов в биомассу или, другими словами, выход биомассы определенного состава из единицы субстрата (углеводов). Эффективность бывает общей и истинной. Общая эффективность включает затраты дыхательного субстрата, связанные с ростом (накоплением биомассы) и с процессами поддержания. При расчете истинной эффективности во внимание принимают только затраты, связанные с ростом растения.
69. Коэффициенты дыхания органов растений картофеля (регрессионный метод)
Орган растения |
Коэффициент дыхания |
|
на рост, г/г |
на поддержание, |
|
|
г/(г • суг) |
|
|
|
|
Листья |
0,46 |
0,04 |
Стебли |
0,25 |
0,03 |
Корни + подземная часть стеблей |
0,19 |
0,03 |
Клубни |
0,04 |
0,01 |
70. Состав сухой биомассы и эффективность роста различных органов растения картофеля (сорт Приекульский ранний)
Орган растения |
Содержание, г/г |
Эффективность роста, г/г |
|||
углеводов |
протеина |
истинная |
общая |
||
|
|||||
Листья |
0,030 |
0,230 |
0,69 |
0,60 |
|
Стебли |
0,060 |
0,115 |
0,80 |
0,70 |
|
Подземная часть стеблей + корни |
0,015 |
0,100 |
0,85 |
0,75 |
|
Клубни: |
|
|
|
|
|
мелкие |
0,410 |
0,065 |
0,96 |
0,90 |
|
крупные |
0,580 |
0,045 |
0,96 |
0,90 |
|
Поскольку энергетические затраты на синтез крахмала примерно в 2 раза ниже по сравнению с затратами на синтез такой же массы белка, то и показатели эффективности превращения ассимилятов в биомассу в клубнях значительно выше, чем в побегах картофеля или плодах других культур, содержащих больше белка.
Эффективность превращения ассимилятов в биомассу закономерно снижается с уменьшением скорости роста у всех органов, особенно быстро, когда относительный прирост биомассы ниже 0,1г/(г ∙сут).
319
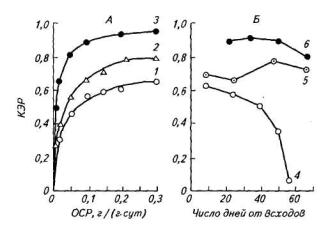
Рис. 103. Коэффициент эффективности роста (КЭР) как функция относительной скорости роста (А) и КЭР (Б) в онтогенезе растений картофеля:
1 — листья; 2—стебли; 3, 6— клубни; 4— надземная часть; 5— целое растение
Выраженный градиент между органами картофеля по эффективности роста и связь этого показателя со скоростью роста дают основания предполагать, что с возрастом растения величина эффективности роста будет изменяться в зависимости от соотношения органов. Из рисунка 103 видно, что формирование клубней не только компенсирует снижение эффективности роста побегов, но и способствует увеличению эффективности роста целого растения.
Эти данные следует учитывать при разработке идеатипа картофеля и в программах селекционного улучшения данной культуры. Полученные результаты указывают на важность оптимизации и контроля за размерами листовой поверхности, так как рост и поддержание донорной функции листьев являются наиболее энергоемкими процессами в растении.
8.6. Минеральное питание
Картофель требователен к условиям произрастания и в первую очередь к почвенному питанию. Концентрации основных минеральных компонентов — N, Р и К в сухой биомассе варьируют в зависимости от органа и возраста растений. Концентрация азота самая высокая (до 6 %) в молодых листьях, калия
— в стеблях и черешках (5—7 %). Содержание фосфора в картофеле на порядок ниже, и органы растения меньше различаются по концентрации этого элемента.
320