

Диссертация на соискание учёной степени
.pdfГЛАВА 1. Обзор литературы
В настоящее время моделированием сердечной активности занимается большое количество исследователей: биологов, биофизиков, физиков-
теоретиков и математиков. Плодотворное сотрудничество учёных, работающих в различных областях знаний, позволило продвинуться в направлении понимания основных механизмов работы сердечных клеток и миокарда в целом. Физико-математическое моделирование в этой области исследований позволяет более четко объяснить многочисленные экспериментальные данные по изучению функции сердца.
1.1 Механизмы сокращения клеток сердечной мышцы
Сердечная мышца относится к возбудимым тканям, клетки которых в ответ на тот или иной раздражитель (электрический, химический, механический)
могут генерировать электрические потенциалы действия – характерные изменения мембранного потенциала клетки, приводящие к ее возбуждению [1, 7]. Более того, некоторые виды клеток могут возбуждаться самопроизвольно. В
основе механизма генерации потенциала действия клетками лежит изменение проницаемости мембран клеток для некоторых ионов (натрия, кальция, калия),
активирующие ионные токи через специальные структуры клеточной мембраны — ионные каналы. Процессы, обеспечивающие сокращение клетки в ответ на электрическую стимуляцию объединяются понятием
электромеханического сопряжения в сердечных клетках. Для мышечного сокращения необходимо повышение внутриклеточной концентрации Са2+ от уровня покоя (около 0.1 мкМ) до 1 мкМ [8].
Сердечные мышечные клетки (кардиомиоциты) включают в себя два основных типа по структуре и функциям: кардиомиоциты рабочего миокарда и клетки водителей сердечного ритма. Первый тип отвечает за механическое сокращение сердца, второй – за формирование устойчивого ритма сердечных сокращений [9].
11

Структура сердечной клетки является достаточно сложной [9, 10]: каждая клетка состоит из нескольких десятков повторяющихся сократительных единиц
– саркомеров, длина которых около 2 мкм; поперечных трубочек (Т-трубочки,
Т-тубулы), представляющих собой глубокие впячивания мембраны,
опоясывают весь кардиомиоцит и локализуются у концов каждого саркомера.
Сокращение саркомера обеспечивается мышечными волоконами
(миофибриллами), расположенными вдоль саркомера и опоясанными сложной сетью внутриклеточных хранилищ ионов кальция, включающих продольный
саркоплазматический ретикулум (СР), терминальные цистерны (ТЦ) (или
люмен), примыкающие к мембране T-тубул (см. рис. 1.1б, рис. 1.3).
а. |
б. |
Рис. 1.1 Структура сердечной клетки. а. Снимок кардиомиоцита, полученный при
помощи электронной микроскопии. б. Трехмерная реконструкция кардиомиоцита.
Обозначения: 1 — сарколемма; 2 — саркоплазматический ретикулум; 3 —
миофибриллы.
На мембране СР в местах контакта СР с Т-тубулами располагается группа специализированных Са2+-высвобождающих каналов, сопряженных с
рианодиновыми рецептороами (RyR-каналы), через которые происходит высвобождение Са2+ из СР. Высвобождение носит триггерный характер, так как открытие RyR-каналов происходит в ответ на приток относительно небольшого количества Са2+ через ионные каналы, расположенных на мембране клетки –
каналы L-типа или дигидропиридиновые рецепторы (см. рис.1.2, 1.3)[8, 10].
12

Рис. |
1.2. |
Схема |
Са2+- |
высвобождающей единицы. |
|
||
На мембране СР RyR-каналы образуют группы (кластеры), состоящие из нескольких сотен каналов. Существуют структуры, состоящие из кластера RyR-
каналов вместе с несколькими L-каналами клеточной мембраны, люменом СР и
диадным пространством между мембранами СР и примыкающими Т-тубулами.
Данные структуры называются Са2+ высвобождающими единицами (Release Unit) (ВЕ, рис.1.2). В кардиомиоците насчитывается несколько десятков тысяч ВЕ; они формируют кальций-высвобождающую систему клетки (рис. 1.3).
Рис. 1.3. Схема кальциевой динамики в сердечной клетке. Схема сопряжения L и RyR-каналов. Адаптировано из
[6]
13
В связи с тем, что высвобождение Са2+ из СР осуществляется через группы
RyR-каналов, исследование их динамических свойств является важной задачей
визучении процессов электромеханического сопряжения.
1.2Рианодиновый рецептор – основной элемент управления кальциевой динамикой в клетке
RyR-канал является гигантским макромолекулярным комплексом (рис. 1.4),
тетрамером, объединяющим четыре субъединицы (полипептидные цепи) с
молекулярной массой 567 кДа каждая [2, 13]. Активность RyR модулируется под действием растительного алкалоида рианодина из коры Ryania speciosa, что и определило его название «рианодиновый рецептор». Существуют три изоформы RyR-каналов: RyR1 (клетки скелетной мускулатуры и мозжечка),
RyR2 (клетки сердечной мышцы и мозга), RyR3 (клетки гладкой мускулатуры).
Они различаются по своей структуре и динамическим параметрам.
Как уже было сказано выше, RyR-каналы располагаются на мембране СР;
при переходе в открытое состояние через них происходит высвобождение ионов Са2+ из просвета ТЦ (люмена). Согласно данным электронной микроскопии [12], RyR-канал имеет форму четырехлистника со стороной 27 нм
(рис. 1.4).
Рис. 1.4. Трехмерная реконструкция
рианодинового канала. а. Вид сбоку. б.
Вид со стороны саркоплазмы. в. Вид
со стороны ТЦ саркоплазматического
ретикулюма.
14

В процессе открытия RyR-канала изменяется его конформационное
(структурное) состояние: данный белок изменяется по форме и структуре,
переходя в проводящее состояние (рис. 1.5).
Рис. 1.5. Открытое (слева) и закрытое (справа) конформационное состояние RyR-канала. Штрихованной линией обозначены трансмембранные сегменты канала, угол между которыми увеличивается при переходе в открытое состояние. Адаптировано из
[7].
На рисунке 1.6 приведены результаты экспериментальных исследований структуры мембраны СР [13]; видно, что на мембранах RyR-каналы объединены в кластеры (решетки) (от 100 до 300 каналов в каждой). Каналы в решетке упорядочены и находятся под определенным углом друг к другу, также экспериментальные данные многих исследовательских групп говорят о существовании взаимодействия между каналами в кластере благодаря наличию между ними специфических связывающих белков [12-15].
а. |
б. |
в. |
Рис.1.6. Кластеры RyR-каналов на мембране СР. (а) Изображение, полученное с помощью электронной микроскопии. (б) Увеличенное и обработанное изображение. (в) Схематическое изображение расположения RyR-канал в кластере.
15
Из всего вышесказанного можно сделать следующие выводы:
RyR-каналы имеют сложную молекулярную структуру и,
соответственно, огромное количество степеней свободы.
На мембране СР каналы образуют регулярную структуру решетки.
Моделирование динамики RyR-каналов усложняется тем, что при разработке теории кластера RyR-каналов необходимо учесть
взаимодействия между ними.
1.3 Эксперименты по изучению изолированных RyR-каналов
Изучая динамические особенности изолированных RyR-каналов независимо
друг от друга несколько авторов исследовало поведение RyR-каналов в лабораторных условиях в липидных бислоях при постоянных концентрациях ионов Са2+ в растворе в условиях отсутствия кальциевой стимуляции [17-19].
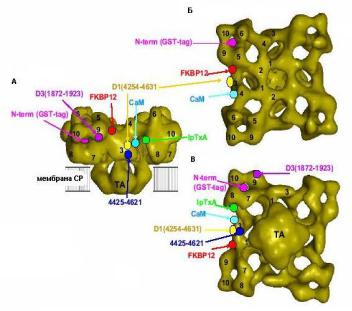
Схема типичной установки представлена на рис. 1.7. В экспериментах раствор разделяется на две части липидным бислоем, имитирующим клеточную мембрану, в котором закрепляется RyR-канал. Та часть, которая соответствует люменальной стороне канала называется trans, цитозольной стороне – cis.
В работах [17-19] помощью подобных установок была проведена серия экспериментов по регистрации ионных токов, проходящих через одиночный канал при различных концентрациях Са2+ в растворе. Отсутствие тока указывало на нахождении канала в закрытом состоянии, а их присутствие – на
его открытие.
Первым важнейшей особенностью RyR-канала как нанообъекта является
стохастический характер процессов открытия/закрытия при постоянных
значениях концентраций и [17-19].
16
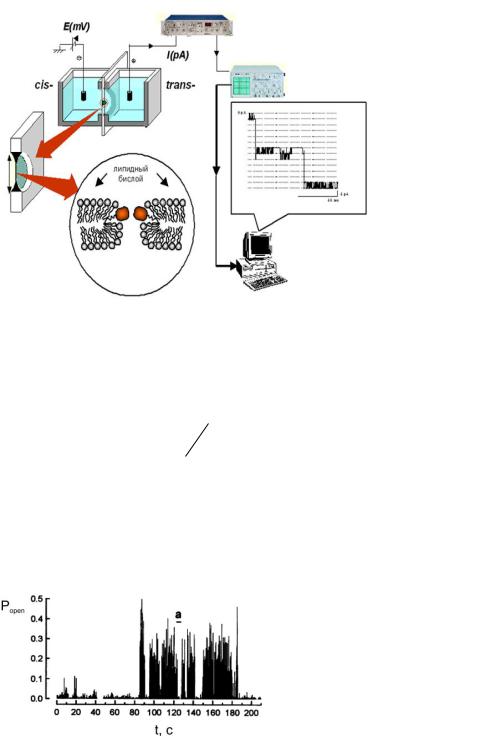
Рис. 1.7. Схема установки по изучению динамики изолированных ионных каналов в липидном бислое. Рисунок адаптирован из: [16].
Важной кинетической характеристикой поведения RyR-канала является вероятность пребывания канала в открытом состоянии ( Popen ), равная отношению суммы времен пребывания в открытом состоянии к длительности
эксперимента: T . При разбиении сигналов на небольшие промежутки времени Т<500 мс в работе [19] исследовалась временная зависимость вероятности пребывания канала в открытом состоянии.
Исследования показали неоднородность функции Popen (t) (рис.1.8).
Рис.1.8. Зависимость вероятности пребывания RyR-канала в открытом состоянии от времени при стационарных условиях.
График зависимости активности канала от времени можно разбить на участки (так называемые пучки (bursts)) с различной вероятностью нахождения канала в открытом состоянии. Данный эффект получил название модовой проводимости (Modal Gating) [21-23]. Так в работе [21] выявлены следующие моды проводимости RyR-канала: H (high Popen ) – мода высокой активности,
17
Popen >0.1, L (low Popen ) – мода низкой активности, 0< Popen <0.1, I (inactivated
mode) – мода нулевой активности, Popen =0 (рис. 1.9).
Рис. 1.9. Ионные токи Са2+ через одиночный RyR-канал в липидном бислое при различных значениях cis[Ca] . Пустыми
прямоугольниками обозначены моды нулевой активности, штрихованными – моды низкой активности, черными – моды высокой активности. Адаптировано из работы [21].
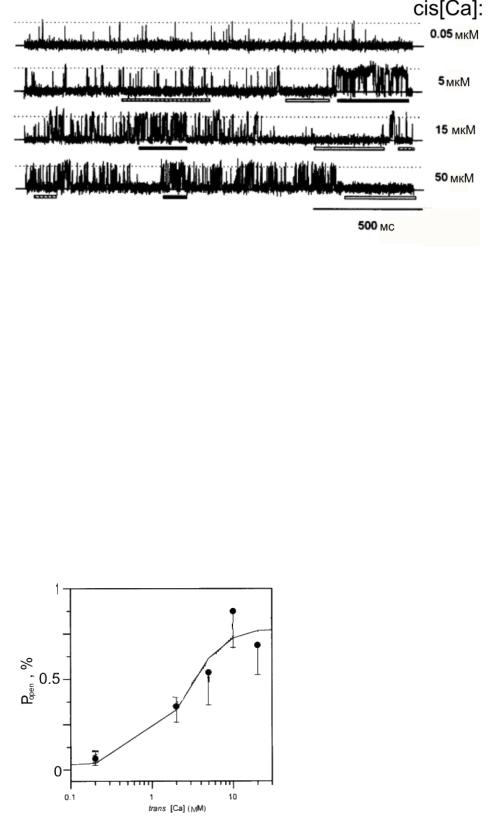
Исследование зависимости вероятности пребывания канала в открытом состоянии от значения концентрации Са2+ в trans-части показало [15, 24, 25],
что в ответ на повышение trans[Ca] увеличивалась частота открытий канала и вероятность Popen (рис. 1.10). Из данного экспериментального факта можно сделать непосредственный вывод, что вероятность открытия RyR-канала увеличивается с ростом концентрации Са2+ в люмене.
Рис. 1.10. Зависимость
вероятности пребывания канала
в открытом состоянии ( Popen ) от
значения trans[Ca] . Рисунок
адаптирован из работы [24].
18
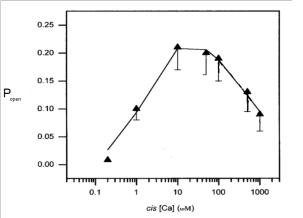
В ряде работ [15, 21, 24], исследовавших влияние концентрации cis[Ca] на активность RyR-канала, показано, что при повышении концентрации Са2+ в
цитозоле (< 10 мкM) наблюдается резкое повышение частоты открытий канала и, соответственно, увеличение вероятности открытия одиночного канала.
Далее, при достижении определенного значения концентрации (> 100 мкM)
происходит спад Popen (cis[Ca]) , то есть наблюдается процесс инактивации RyR-
канала при повышенной концентрации цитозольного кальция (рис.1.11).
Рис. 1.11. Зависимость вероятности пребывания канала в открытом состоянии от значения cis[Ca] .
Рисунок адаптирован из работы Gyorke S. [20]
Уменьшение значения Popen при высоких значениях cis[Ca] объясняется многими исследователями [12, 15, 21, 26] тем фактом, что, являясь Са2+-
активируемым рецептором, RyR-канал имеет на cis-стороне два типа активных центров, способных связаться с ионами Са2+: активационный и инактивационный [26-28]. На рисунке 1.12 изображена классическая схема активации/инактивации канала [27].
19

Рис. 1.12. Диаграмма связывания ионов Са2+ с активационным (1) и инактивационным центрами канала. Из покоящегося (закрытого) состояния в активированное (открытое) RyR-канал может вывести быстрое связывание Са2+ с активационным центром. Процессы связывания с инактивационным центром, имеющим более высокое сродство к Са2+, считаются медленными относительно процессов активации.
Сложная регуляция RyR-канала кальцием впервые была обнаружена в работе [29] при проведении экспериментов в плоском липидном бислое.
Показано, что резкое увеличение cis[Ca] (~ 1 мс) сначала быстро активирует одиночный RyR-канал до высокого уровня вероятности Popen , а затем значение
Popen медленно уменьшается со временем при фиксированной концентрации Са2+ (рис. 1.13). Этот эффект получил название адаптации канала к продолжительной стимуляции.
Рис. 1.13. Эффект адаптации RyR-канала к продолжительной стимуляции. а. Временная зависимость вероятности Popen
при ступенчатом увеличении уровня cis[Ca] (б.) Рисунок адаптирован из работы [25].
Следует отметить, что в экспериментальных работах повышение уровня cis[Ca] проводилось двумя способами: резкое повышение с помощью лазерного флэш-фотолиза [29-33] и постепенное повышение
20