

Диссертация на соискание учёной степени
.pdf
|
40 |
|
|
|
|
|
|
35 |
|
|
|
|
|
|
30 |
|
|
|
|
|
событий |
25 |
|
|
|
|
|
20 |
|
|
|
|
|
|
|
|
|
|
|
|
|
Число |
15 |
|
|
|
|
|
10 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
5 |
|
|
|
|
|
|
0 |
|
|
|
|
|
|
0.00 |
0.01 |
0.02 |
0.03 |
0.04 |
0.05 |
Popen
Рис. 3.28. Распределение по вероятности пребывания в открытом состоянии.
3.7.2 Зависимость активности RyR-канала от уровня cis[Ca]
На рисунке 3.29 изображены зависимости активности RyR-канала от времени при различных значениях cis[Ca] (открытое состояние соответствует значению 1).
Рис. 3.29. Активность RyRканала при стационарных услових для трех различных значений концентрации цитозольного кальция cis[Ca]. Нулю соответствует закрытое состояние.
Мода H выделена черными сплошными линиями, мода L – пунктирными линиями. Белыми сплошными линиями обозначена мода нулевой активности канала
I.
Для сравнения результатов численных реализаций с экспериментальными данными, на рисунке 3.30 представлены графики экспериментальные регистрации активности RyR-канала при различных значениях cis[Ca] [15].
111

а. |
б. |
Рис. 3.30. Активность RyR-канала при стационарных услових при трех различных значениях концентрации цитозольного кальция cis[Ca]. а. Результаты численных экспериментов. б. Результаты экспериментов, адаптированные из работы [15]. Нулю соответствует закрытое состояние. Участки с высокой вероятностью Popen обозначены черными сплошными линиями, с малой –
пунктирными линиями, длительные пребывания в закрытом состоянии – белыми сплошными линиями.
Сравнивая рисунок 3.30а с результатами моделирования (рис. 3.30б), можно сделать вывод, что результаты численных экспериментов находятся в хорошем согласии с экспериментальными данными.
В работе при проведении анализа кинетических характеристик динамики
RyR-канала в рамках модели, были построены графики зависимости Popen от концентрации Са2+ в cis-части при различных значениях числа кластеров в активных центрах канала, представленные на рисунке 3.31.
112

а. |
|
|
|
|
|
|
Рис. |
3.31. |
|
Влияние |
|||
|
70 |
z = 20 |
|
|
|
|
цитозольного |
|
Са2+ |
на |
|||
|
|
|
|
|
|
активность |
RyR-канала. |
а. |
|||||
|
60 |
z = 25 |
|
|
|
|
|||||||
|
z = 30 |
|
|
|
|
Зависимости |
|
пребывания |
|||||
|
|
|
|
|
|
|
|||||||
|
|
|
|
|
|
|
|
||||||
|
50 |
|
|
|
|
|
канала в открытом состоянии |
||||||
|
|
|
|
|
|
|
|||||||
% |
40 |
|
|
|
|
|
от |
cis[Ca] |
при |
различных |
|||
, |
|
|
|
|
|
|
значениях |
|
|
количества |
|||
open |
30 |
|
|
|
|
|
|
|
|||||
|
|
|
|
|
|
|
|
|
|
|
|
||
P |
|
|
|
|
|
секторов |
активных |
центров |
|||||
|
|
|
|
|
|
||||||||
|
|
|
|
|
|
|
|||||||
|
20 |
|
|
|
|
|
RyR-канала. |
|
|
|
|
||
|
|
|
|
|
|
|
|
|
|
|
|||
|
10 |
|
|
|
|
|
б. |
Зависимости |
вероятности |
||||
|
0 |
|
|
|
|
|
пребывания |
|
|
|
в |
||
|
|
|
|
|
|
инактивированном |
состо- |
||||||
|
|
0.1 |
1 |
10 |
100 |
1000 |
|||||||
|
|
|
cis[Ca], мкM |
|
|
янии от времени при трех |
|||||||
|
|
|
|
|
|
|
|||||||
б. |
|
|
|
|
|
|
значениях |
числа |
секторов |
в |
|||
|
80 |
|
|
|
|
|
активных центрах канала. |
|
|||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
70 |
|
|
|
|
|
|
|
|
|
|
|
|
|
60 |
|
|
|
|
|
|
|
|
|
|
|
|
, % |
50 |
|
z = 20 |
|
|
|
|
|
|
|
|
|
|
|
z = 25 |
|
|
|
|
|
|
|
|
|
|
||
|
|
|
|
|
|
|
|
|
|
|
|
||
inact |
40 |
|
z = 30 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||
P |
30 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
20 |
|
|
|
|
|
|
|
|
|
|
|
|
|
10 |
|
|
|
|
|
|
|
|
|
|
|
|
|
0 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
0.1 |
1 |
10 |
100 |
1000 |
|
|
|
|
|
|
|
|
|
|
cis[Ca], мкМ |
|
|
|
|
|
|
|
|
|
|
Зависимость активности RyR-канала при стационарных условиях от
концентрации |
цитозольного Са2+ носит колоколообразный характер: |
Popen (cis[Ca]) |
растет с увеличением cis[Ca] в интервале от 0 до 100 мкМ и |
спадает при cis[Ca]>100 мкМ.
Эффект понижения Popen при достаточно больших значениях cis[Ca] получил в литературе название Са2+-зависимой инактивации [24] и является одним из ключевых феноменов тонкой регуляции RyR-канала ионами Са2+. Однако следует отметить, что концентрации cis[Ca]>100 мкМ не являются физиологическими и представляют интерес только для экспериментов in vitro.
Приведенные на рисунке 3.31б графики зависимости вероятности пребывания в инактивационном состоянии ( Pinact ) от cis[Ca] говорят о том, что с ростом cis[Ca] увеличивается частота электронных переходов в инактивационное состояние, в связи с этим повышается Pinact .
113
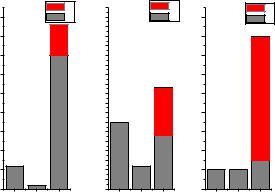
При исследовании динамических рядов активности RyR-канала при различных значениях cis[Ca] были изучены моды активности канала и построена гистограмма распределения по пребываниям в той или иной моде,
представленная на рисунке 3.32.
|
|
I |
90 |
|
C |
80 |
|
|
70 |
|
|
60 |
|
|
50 |
|
|
40 |
|
|
30 |
|
|
20 |
|
|
10 |
|
|
0 |
|
|
H |
L |
C/I |
cis[Ca]=0.5 мкМ |
||
95 |
|
I |
90 |
|
C |
85 |
|
|
80 |
|
|
75 |
|
|
70 |
|
|
65 |
|
|
60 |
|
|
55 |
|
|
50 |
|
|
45 |
|
|
40 |
|
|
35 |
|
|
30 |
|
|
25 |
|
|
20 |
|
|
15 |
|
|
10 |
|
|
5 |
|
|
0 |
|
|
H |
L |
C/I |
cis[Ca]=100 мкМ |
||
|
|
I |
90 |
|
C |
80 |
|
|
70 |
|
|
60 |
|
|
50 |
|
|
40 |
|
|
30 |
|
|
20 |
|
|
10 |
|
|
0 |
|
|
H |
L |
C/I |
cis[Ca]=1000 мкМ |
||
Рис. 3.32. Гистограммы распределений по пребываниям в модах H, L и C/I. Первый столбец – мода H, второй – мода L, третий – мода C/I. Верхняя часть третьего столбца – мода I, нижняя – мода С.
Были введены следующие моды активности:
Мода С/I соответствует пребыванию канала в закрытом состоянии С или в инактивационном состоянии I. На гистограмме состояния С и I разделены в третьем столбце. Вероятность пребывания в открытом состоянии в этой моде является нулевой.
Моде H соответствуют длительные пребывания в открытом состоянии О и частые переходы из закрытого состояния в открытое
(при этом время пребывания в открытом состоянии больше, чем в закрытом: topen tclosed ). В данной моде Popen
Моде L соответствует серия кратковременных пребываний в открытом состоянии ( topen tclosed ). Закрытия происходят вследствие переходов в конформационно закрытое состояние и в инактивацинное состояние. В моде низкой активности:
0 P |
P . |
open |
m |
Как ясно из рисунков 3.31 и 3.32, с ростом cis[Ca] наблюдается увеличение вероятности нахождения системы в инактивационном состоянии, и оно
114
становится превалирующим после значения cis[Ca]=100 мкМ, которое соответствует максимуму зависимости Popen (cis[Ca]) .
Согласно экспериментальным данным [20], активность RyR-каналов в кардиомиоцитах становится нулевой при очень высоких значениях концентрации цитозольного Са2+ (5-10 мМ), которые, предположительно,
недостижимы в клетках. Однако, Са2+-зависимая инактивация также как и адаптация, является механизмом негативного контроля, обеспечивающим своевременное завершение процесса высвобождения Са2+ из СР во время клеточного цикла.
3.7.3 Влияние ионов Mg2+ на динамику одиночного RyR-канала
Ионы Mg2+ как и ионы Са2+ обладают сильным сродством с активационной частью RyR-канала, однако, занимая вакантные места на активационных частях, они не приводят к открытию канала [122, 123].
Как и в случае взаимодействия ионов Са2+с активными центрами, в
настоящей диссертационной работе предлагается статистический подход к рассмотрению конкурентного присоединения ионов Mg2+ и Са2+ к активным центрам канала.
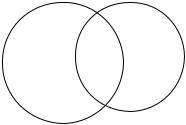
На рисунке 3.33а схематично представлен процесс присоединения ионов Са2+ и Mg2+ с активному центру канала. Заполнение ионами активных мест связывания активного центра описано с помощью диаграммы Эйлера (рис. 2.31б).
115
а. |
|
|
|
|
|
|
|
б. |
|
||
2+ |
|
Ca2+/ |
2+ |
2+ |
|
|
|
||||
|
|
|
|
|
|
|
|
||||
|
|
|
|
|
|
|
|
||||
|
Ca |
|
Mg2+ |
Mg |
Ca |
|
|
|
|
|
|
|
2+ |
2+ |
|
Ca2+/ |
2+ |
|
|
|
|
|
|
|
Mg |
Ca |
|
Mg2+ |
Mg |
|
|
|
|
|
Ca2+/ Mg2+ |
|
|
|
|
|
|
Ca2+ |
|||||
|
|
|
|
|
|
|
|
|
|
||
|
|
|
|
|
|
|
|
||||
|
|
2+ |
|
2+ |
2+ |
|
|
|
|
|
Mg2+ |
|
|
Mg |
|
Ca |
Ca |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
2+ |
Ca2+/ |
Ca2+/ |
2+ |
|
|
2+ |
|
|
|
|
|
Mg |
Mg2+ |
Mg2+ |
Mg |
|
Mg |
|
|
|
Свободные активные части |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Ca2+ |
|
|
Ca2+ |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||
|
|
|
|
|
|
|
|
|
|
||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
2+ |
|
Ca2+/ |
|
|
2+ |
|
|
|
|
|
|
Mg |
|
Mg2+ |
|
Mg |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Рис. 3.33 Присоединение ионов Са2+ и Mg2+ к активным центрам. а. Схема активных центров RyR-канала, занятых ионами Са2+, Mg2+, либо двумя типами ионов. б. Диаграмма Эйлера, показывающая заселенность кластеров активных центров канала.
Было сделано предположение, что если в кластер активного центра попадают оба типа ионов, то вероятность присоединения того или иного типа ионов к свободному центру равняется ½.
Пусть m |
[Mg2 ] |
|
|
|
2+ |
|
|
|
|
– относительная концентрация ионов Mg |
вблизи канала, |
||||
[Mg2 ] |
|||||||
|
max |
|
|
|
|
|
|
где [Mgmax2 ] – максимальная концентрация магния, при которой заполнены все |
|||||||
места связывания |
активного центра канала; |
c |
cis[Ca] |
– |
относительная |
||
|
|||||||
cis[Ca] |
|||||||
|
|
|
|
|
max |
|
|
концентрация ионов Са2+. |
|
|
|
|
|||
|
Далее определялась вероятность нахождения более чем k ионов Са2+ в |
|||||||
активном центре канала с учётом присутствия на нем ионов Mg2+. |
|
|||||||
|
Относительное число активных мест, в которых находятся только ионы Са2+ |
|||||||
в кластере: PCa c(1 m) ; |
относительное число секторов, в которых находятся |
|||||||
ионы Са и Mg, но присоединяются ионы Са: P |
|
1 c m |
. Соответственно, |
|||||
|
|
|
|
Ca/Mg Ca |
2 |
|
|
|
|
|
|
|
|
|
|
|
|
относительное |
число |
«сработавших» |
ионов |
Са2+ |
равно |
|||
P |
|
c(1 m) 1 cm c(1 1 m) . Число комбинаций k ионов Са2+ |
в z частях |
|||||
Ca total |
|
2 |
2 |
|
|
|
|
|
|
|
|
|
|
|
|
||
116
кластера равно Czk |
z! |
. Наконец, вероятность обнаружения более чем k |
||
|
|
|||
k!(z k)! |
||||
|
|
|||
ионов Са2+ в кластере активной части равняется:
P (k, z,c) |
z |
C |
|
c(1 m / 2) |
|
1 c(1 m / 2) |
|
z . |
(3.21) |
Ca |
z |
|
|
|
|
|
|||
|
k |
|
|
|
|
|
|
|
|
В численных экспериментах значение cis[Ca] варьировалось в диапазоне от
0.05 до 1000 мкМ. В первой части экспериментов не учитывалось влияние ионов Mg2+, и интенсивность электронных переходов в зависимости от cis[Ca]
z |
z! |
|
|
|
||
рассчитывалась по формуле (c) Pkz (c) / dt |
c (1 |
c)z / dt |
(см. |
|||
|
|
|||||
!(z )! |
||||||
k |
|
|
|
|||
формулу 2.10), где с – относительная концентрация ионов Са2+ в cis-части.
Во второй серии экспериментов учитывалось присутствие Mg2+ в растворе
(Mg=3 мМ), и a b (cis[Ca]) и i b (cis[Ca]) рассчитывались согласно формуле
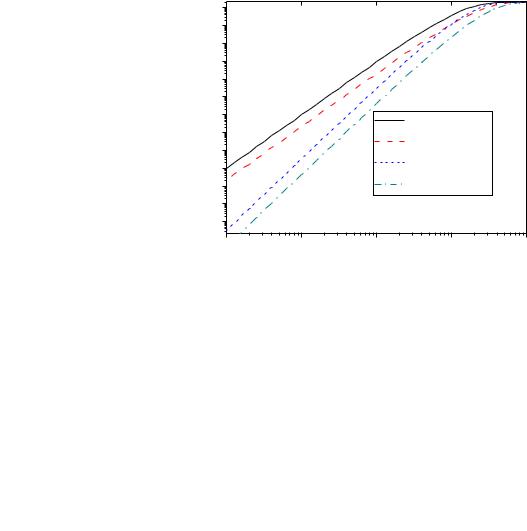
(3.21). Графики зависимостей интенсивностей электронных переходов с учетом и без учета Mg2+ представлены на рисунке 3.33.
|
|
10 |
|
|
|
|
|
|
1 |
|
|
|
|
|
|
0.1 |
|
|
|
|
|
|
0.01 |
|
|
|
|
|
|
1E-3 |
|
|
|
|
|
|
1E-4 |
|
|
|
|
elect |
1E-5 |
|
|
a b(Mg=0) |
|
|
1E-6 |
|
|
|
|||
|
|
|
|
a b(Mg=3 мМ) |
|
|
|
|
1E-7 |
|
|
|
|
|
|
1E-8 |
|
|
i b(Mg=0) |
|
|
|
1E-9 |
|
|
i b(Mg=3 мМ) |
|
|
1E-10 |
|
|
|
|
|
|
1E-11 |
|
|
|
|
|
|
|
0.1 |
1 |
10 |
100 |
1000 |
|
|
|
|
cis[Ca], мкМ |
|
|
Рис. 3.34 Зависимости интенсивностей электронных переходов канала от уровня cis[Ca] с учетом ионов Mg2+ и без учета. Параметры модели для активационного центра: k = 4, z=25. Для инактивационного центра: k = 7, z=25 (см. п. 2.1.8).
117
Как видно из рисунка, учет Mg2+ сдвигает зависимость |
elect |
(cis[Ca]) вниз, то |
|||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
есть статистический подход хорошо описывает ингибирующие свойства ионов |
|||||||||||||
Mg2+. |
|
|
|
|
|
|
|
|
|
|
|
|
|
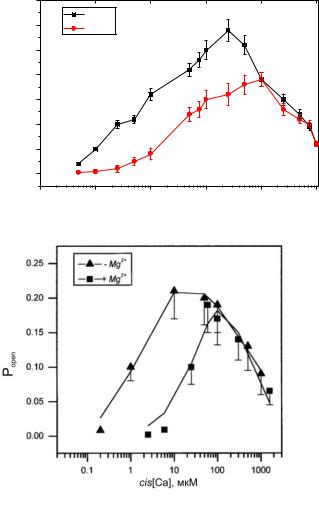
На рисунке 3.35 представлены графики зависимости вероятности |
|||||||||||||
пребывания канала в открытом состоянии от концентрации Са2+ в cis-части в |
|||||||||||||
отсутствии и в присутствии ионов Mg2+ P |
|
(cis[Ca]) . |
|
|
|
|
|||||||
|
|
|
|
|
|
open |
|
|
|
|
|
|
|
а. |
|
|
|
|
|
|
|
Рис. 3.35. а. |
Зависимость |
||||
|
0.35 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
- Mg2+ |
|
|
|
|
вероятности |
|
|
|
Popen |
||
|
0.30 |
+ Mg |
2+ |
|
|
|
|
|
|
|
|||
|
|
|
|
|
|
|
|
|
|
|
|||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
0.25 |
|
|
|
|
|
|
обнаружения |
|
RyR-канала |
|||
open |
0.20 |
|
|
|
|
|
|
в открытом состоянии от |
|||||
|
|
|
|
|
|
|
|||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
P |
0.15 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
цитозольной |
|
|
|
|
||
|
|
|
|
|
|
|
|
|
|
|
|
||
|
0.10 |
|
|
|
|
|
|
концентрации |
|
|
кальция |
||
|
|
|
|
|
|
|
|
|
|
||||
|
0.05 |
|
|
|
|
|
|
( cis[Ca]), построенная по |
|||||
|
|
|
|
|
|
|
|
||||||
|
0.00 |
|
|
|
|
|
|
|
|
|
|
|
|
|
0.01 |
0.1 |
|
1 |
10 |
100 |
1000 данным |
|
численных |
||||
|
|
|
|
|
cis[Ca], мкM |
|
|
экспериментов |
на |
ЭК- |
|||
|
|
|
|
|
|
|
|
||||||
б. |
|
|
|
|
|
|
|
модели RyR-канала в |
|||||
|
|
|
|
|
|
|
|
||||||
|
|
|
|
|
|
|
|
отсутствие ионов Mg2+ и с |
|||||
|
|
|
|
|
|
|
|
учетом Mg2+ (3 мМ). |
|
||||
|
|
|
|
|
|
|
|
б. |
Экспериментальная |
||||
|
|
|
|
|
|
|
|
зависимость |
|
вероятности |
|||
|
|
|
|
|
|
|
|
обнаружения |
|
RyR-канала |
|||
|
|
|
|
|
|
|
|
в открытом состоянии от |
|||||
|
|
|
|
|
|
|
|
цитозольной |
|
|
|
|
|
|
|
|
|
|
|
|
|
концентрации |
|
|
кальция |
||
|
|
|
|
|
|
|
|
( cis[Ca]). |
|
|
Рисунок |
||
|
|
|
|
|
|
|
|
адаптирован |
|
из |
работы |
||
|
|
|
|
|
|
|
|
[20]. |
|
|
|
|
|
Как видно из графика, зависимости имеют колоколообразную форму с |
|||||||||||||
максимумом, соответствующим значению |
|
cis[Ca]=50 мкМ (без учета Mg2+) и |
|||||||||||
cis[Ca]=100 мкМ (с учетом Mg2+). Другими словами, при учете ионов Mg2+ |
|||||||||||||
происходит сдвиг вправо зависимости P |
(cis[Ca]) . |
Для сравнения на рисунке |
|||||||||||
|
|
|
|
|
|
open |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
118 |
3.35б приведен график из работы [20]. Анализ представленных кривых говорит
о том, что результаты моделирования с высокой степенью точности совпадают
сэкспериментальными данными.
Вприсутствии ионов Mg2+ в растворе, в котором находится RyR-канал,
график зависимости Popen (cis[Ca]) сдвигается вниз и вправо, то есть максимум достигается при больших значениях cis[Ca] и максимум функции меньше, чем в отсутствие ионов Mg2+, конкурирующих с активными ионами Са2+.
Ионы Mg2+ являются потенциальными ингибиторами RyR-каналов, в связи с этим находят активное применение в фармакологии и терапии для понижения частоты высвобождений Са2+ для предотвращения аритмии и других патологий.
Модификация электронно-конформационной модели и введение инактивационного состояния позволили описать не только основные свойства изолированных RyR-каналов, но и важнейшие тонкие эффекты взаимодействия каналов с ионами Са2+, которые являются механизмами негативного контроля и должны учитываться при моделировании процесса КВВК.
3.8Заключение
Вданной главе приведены результаты параметрического анализа
электронно-конформационной модели одиночного RyR-канала, сравнения
результатов |
ЭК-модели |
с традиционными теориями, |
основанными |
на |
марковских цепях. |
|
|
|
|
В рамках |
ЭК-модели |
проведена серия численных |
экспериментов |
по |
исследованию:
процессов открытия RyR-канала при резком повышении уровня cis[Ca];
процесса закрытия RyR-канала при резком уменьшении уровня cis[Ca];
процесса адаптации RyR-канала к установившемуся уровню cis[Ca];
зависимости вероятности пребывания RyR-канала в открытом и закрытом состояниях при постоянном уровне cis[Ca];
влияния ионов Mg2+ на активность RyR-канала при стационарных условиях.
119
зависимости конформационной координаты Q от времени методом Херста.
Сравнительный анализ показал, что результаты проведенных численных экспериментов хорошо согласуются с известными экспериментальными данными, и ЭК-модель дает адекватное описание стохастической динамики
RyR-канала.
120