

Диссертация на соискание учёной степени
.pdfа. 1.0 |
|
|
a u = 0.1 |
-1 |
|||
|
0.8 |
tun |
= 0 |
мс |
|||
|
a u = 0.2 |
мс-1 |
|||||
open0.6 |
|
|
|||||
P |
0.4 |
|
|
|
a u |
= 1 мс-1 |
|
|
|
|
|
|
|
|
|
|
0.2 |
|
|
|
|
|
|
|
0.0 |
|
|
|
|
|
|
б. |
0 |
10 |
|
20 |
30 |
|
|
|
|
|
|
1.0 |
|
tun = 0.02 мс-1 |
|
|
|
0.8 |
|
|
|
||
open 0.6 |
|
|
|
|
|
P |
|
|
|
|
|
0.4 |
|
|
|
|
|
0.2 |
|
|
|
|
|
0.0 |
|
|
|
|
|
в. |
0 |
10 |
|
20 |
30 |
1 |
|
|
|
|
|
cis[Ca],мкМ |
|
|
|
|
|
0 |
|
|
|
|
|
|
0 |
10 |
t, мс |
20 |
30 |
Рис. 3.20. Зависимость вероятности пребывания канала в открытом |
|
||||
состоянии от времени в отсутствии туннелирования (а) и при учете |
|
||||
туннелирования (б) после резкого понижения уровня cis[Ca] (в). |
|
||||
Видно, что зависимость P (t) носит экспоненциальный характер, причем
open
по мере возрастания a u увеличивается скорость спадания Popen . Из графика этой зависимости можно определить постоянную времени закрытия closing , ей соответствует момент времени t, при котором Popen уменьшается в е раз и
достигает уровня |
1 |
0.368 (пунктирная линия на рисунке). |
|
|||||||
|
|
e |
|
|
|
|
|
|
|
|
На рисунке 3.21 изображена зависимость среднего времени закрытия канала |
||||||||||
closing |
от интенсивности закрытий канала a u . |
|
|
|
|
|
||||
|
40 |
|
|
|
Рис. |
3.21. |
Зависимость |
|||
|
35 |
|
|
tun = 0 |
среднего времени закрытия |
|||||
|
|
|
|
одиночного RyR-канала от |
||||||
|
30 |
|
|
tun = 0.02 мс-1 |
||||||
|
|
|
интенсивности |
переходов, |
||||||
мс |
25 |
|
|
|
|
|
|
|
|
|
|
|
|
|
связанных с отсоединением |
||||||
, |
20 |
|
|
|
|
|
|
|
|
|
closing |
|
|
|
ионов |
|
Са2+ |
от |
активных |
||
|
|
|
|
|
||||||
15 |
|
|
|
|
|
|
|
|
|
|
t |
|
|
|
частей канала в отсутствии |
||||||
|
|
|
|
|||||||
|
|
|
|
|
||||||
|
10 |
|
|
|
туннельных |
переходов |
и |
|||
|
|
|
|
|
||||||
|
5 |
|
|
|
при |
tun |
0.02 мс 1 . |
|
||
|
|
|
|
|
|
|
|
|
|
|
|
0 |
|
|
|
|
|
|
|
|
|
|
|
|
0.1 |
1 |
|
|
|
|
|
|
|
|
|
|
, мс-1 |
|
|
|
|
|
|
|
|
|
|
a u |
|
|
|
|
|
|
101

Как видно из рисунка, величина closing падает с увеличением a u , при этом,
с учетом процесса туннелирования в эксперименте канал закрывается быстрее.
Этот эксперимент проводился на малых интервалах времени (t = 15 мс).
Пренебрегая процессами переходов в инактивационное состояние, процесс
закрытия канала можно описать с помощью простейшей марковской схемы:
O |
a u |
C , |
(3.18) |
a b tun |
где C – закрытое состояние, O – открытое состояние.
Рассмотрим случай при tun =0. Вероятности пребывания канала в состояниях О и С описываются системой дифференциальных уравнений Колмогорова:
dPC (t) a u PO (t) a b PC (t)
dt (3.19)
dPO (t) a b PC (t) a u PO (t), dt
где PC (t) и PO (t) – вероятности пребывания канала в состояниях О и С,
соответственно.
В начальный момент времени все каналы находятся в открытом состоянии,
поэтому начальные условия для уравнений (3.19) следующие: PC (t 0) 0 ,
PO (t 0) 1.
С учетом этих начальных условий решение системы (3.19) выглядит как:
PC |
(t) |
a u a ue |
( a b u b ) t |
|
|
||
|
|
|
|
||||
|
a u a b |
|
|
||||
|
|
|
. |
(3.20) |
|||
|
|
a ue |
(Λa b +Λu b ) t |
a b |
|||
|
(t) |
|
|
||||
PO |
|
|
|
|
|||
|
Λa u +Λa b |
|
|
|
|||
|
|
|
|
|
|
||
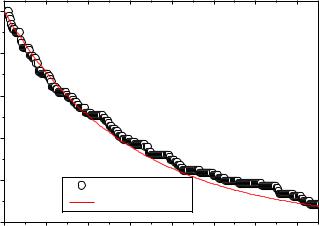
На рисунке 3.22 представлены графики зависимости Popen (t) при a u =0.2
мс-1 ( dt 0.05 мс, a b =0.2 мс-1), полученные по результатам численных экспериментов, а также кривая, соответствующая аналитическому решению уравнений Колмогорова (3.20).
102
|
1.0 |
|
|
|
|
|
|
|
|
0.8 |
|
|
|
|
|
|
|
open |
0.6 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
P |
0.4 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
0.2 |
|
|
Popen(эксперимент) |
|
|
|
|
|
|
|
|
|
|
|
||
|
|
|
|
Popen=100*exp(-t/5) |
|
|
|
|
|
0.0 |
|
|
|
|
|
|
|
|
0 |
2 |
4 |
6 |
8 |
10 |
12 |
14 |
t, мс
Рис. 3.22. Зависимость вероятности нахождения канала в открытом состоянии от времени после резкого понижения cis[Ca] и решение уравнения (3.19) при
a u 0.2 мс 1 и a b 0 .
Как видно из графика, результаты численных экспериментов хорошо согласуются с простейшим марковским приближением.
Экспериментальные данные по изучению закрытия RyR-каналов указывают,
что среднее время закрытия имеет порядок 5 мс [119], что также хорошо согласуется с данными численных экспериментов, представленных на рисунке
3.22.
Согласно экспериментальным данным отношение постоянной времени закрытия канала к постоянной времени открытия closing / opening ~5, в связи с
этим в терминах ЭК-модели можно сделать вывод, что вероятность электронных переходов, связанных с присоединением Са2+ к активационному центру выше вероятности переходов, связанных с отсоединением Са2+ от активационного центра ( Pa b Pa u ).
3.6 Процесс адаптации RyR-каналов к продолжительной стимуляции
Далее в данной работе проведено моделирование динамики одиночных
RyR-каналов, а именно, изучение поведения каналов при продолжительной
стимуляции.
103

Впервые явление медленного понижения активности канала при длительном стимуле было обнаружено экспериментально в 1993 году [25] и получило название адаптации RyR-канала к продолжительной стимуляции. Результаты исследований из работы [25] представлены на рисунке 3.23б. Подробно этот эффект описан ранее в первой главе настоящей работы.
Врамках данной работы исследована вероятность пребывания канала в открытом состоянии при учете всех видов электронных переходов, включая переходы в инактивированное состояние, на больших интервалах времени (t > 3 c).
Также как и в экспериментах, описанных выше, исследовалась динамика ансамбля независимых RyR-каналов, в начальный момент времени все каналы находились в закрытом состоянии.
Вмомент времени t=0 уровень cis[Ca] резко повышается до 1 мкМ, при этом, вероятности электронных переходов становятся больше нуля.
На рисунке 3.23 представлены графики зависимости Popen и cis[Ca] от
времени.
а. |
|
|
|
|
|
б. |
|
1.0 |
|
|
|
|
|
open |
I |
II |
|
|
|
|
|
|
|
|
|
||
P |
0.5 |
|
III |
|
|
|
|
|
|
|
|||
|
|
|
|
|
||
|
|
|
|
|
|
|
мкМ |
0.0 |
|
|
|
|
|
1 |
0 |
1 |
2 t, c 3 |
4 |
5 |
|
cis[Ca], |
0 |
|||||
|
|
|
|
|
|
|
Рис. 3.23. Зависимости вероятности Popen пребывания канала в открытом
состоянии при ступенчатом изменении cis[Ca] от времени. а. Результаты численного эксперимента. б. Экспериментальные данные, адаптированные из
[25].
Условно график зависимости P (t) можно разбить на три участка (рис.
open
3.23а). На первом из них (0<t<100 мс) происходит активация каналов в
ансамбле, |
то есть резкое повышение Popen до 0.85 с постоянной времени |
open 2.5 |
мс (участок I). Подробно процесс активации рассмотрен в разделе |
3.4. |
|
104
Далее в интервале 100 мс < t < 2.5 с наблюдается медленный спад Popen , до
стационарного значения P 0.1 (участок II). После этого система приходит в
open
динамическое равновесие (участок III).
На больших интервалах времени (~3c) можно пренебречь медленной конформационной динамикой, поэтому на участке II переходы RyR-канала из одного состояния в другое можно описать с помощью марковского процесса
(3.10). Уравнения (3.11) имеют следующие начальные условия:
P (t 0) 1, P (t 0) 0, P (t 0) 0, P (t 0) 0 . |
|||
C |
O |
I1 |
I2 |
Точные решения этих уравнений (аналитический вид здесь не приводится)
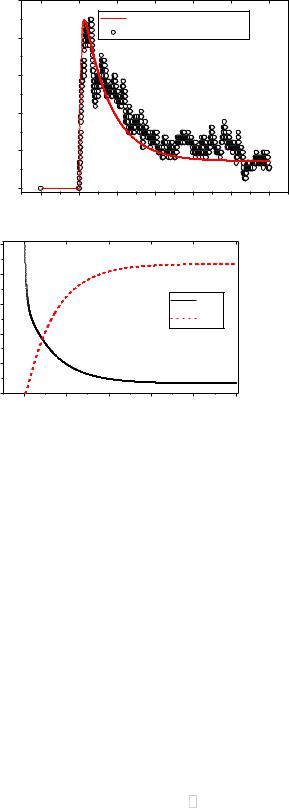
представлены в виде графиков на рисунке 3.24а, 3.24б. Для сравнения с точным решением (сплошная линия) на рисунке 3.24а также приведены данные численных экспериментов. Эти результаты изображены ранее на рисунке 3.23а.
105
а. |
|
1.0 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Марковское приближение |
|
||
|
|
0.8 |
|
|
Численный эксперимент |
|
||
|
|
|
|
|
|
|
|
|
|
|
0.6 |
|
|
|
|
|
|
|
open |
|
|
|
|
|
|
|
P |
0.4 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
0.2 |
|
|
|
|
|
|
|
|
0.0 |
|
|
|
|
|
|
|
|
-1 |
0 |
1 |
2 |
3 |
4 |
5 |
|
|
|
|
|
t, с |
|
|
|
б. |
|
|
|
|
|
|
|
|
100 |
|
|
|
|
|
|
|
|
|
80 |
|
|
|
|
|
|
|
|
60 |
|
|
|
|
|
P |
|
% |
|
|
|
|
|
|
close |
|
|
|
|
|
|
|
Pinact |
|
|
P, |
|
|
|
|
|
|
|
|
40 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
20 |
|
|
|
|
|
|
|
|
0 |
|
|
|
|
|
|
|
|
|
0 |
1 |
2 |
3 |
4 |
5 |
|
t, c
Рис. 3.24. Эффект адаптации RyR-канала к продолжительной стимуляции а. Зависимости вероятностей нахождения канала
воткрытом состоянии от времени, полученные из численных экспериментов и из точного решения уравнений Колмогорова. б. Временные зависимости нахождения канала
взакрытом и инактивированном состоянии, полученные из уравнений (3.11). Интенсивности электронных переходов при
t>0: |
|
a b |
0.01 мс 1 |
, |
|
a u |
0.01 |
мс-1, |
|
i b |
0.002 мс 1 |
, |
||
|
|
|
|
|
|
|
|
|
|
|
|
|||
|
i u |
0.0002 мс 1 . |
tun |
0.002 мс 1 . |
|
|
|
|
|
|||||
|
|
|
|
|
|
|
|
|
|
|
|
|||
Из анализа рисунка 3.24а можно сделать вывод, что результаты моделирования хорошо согласуются с точными решениями уравнений
Колмогорова в марковском приближении. |
|
|
|
||
Вероятность |
P (t) |
сначала после |
достижения пика |
уменьшается |
|
|
open |
|
|
|
|
экспоненциально, |
затем при t 3 c вероятности Popen, Pclose |
и Pinact выходят на |
|||
стационарный уровень. |
Уменьшение P |
(t) при t<2.5 |
с |
указывает на |
|
|
|
open |
|
|
|
медленный спад активности канала на этом промежутке времени. При анализе данных численных экспериментов было выявлено, что стационарный уровень
106
P (t) проявляется в связи со стремлением канала к динамическому
open
равновесию между переходами в состояния: O, C и I, что подтверждает выход на стационарный уровень вероятностей переходов в состояние О и I (рис.
3.24б).
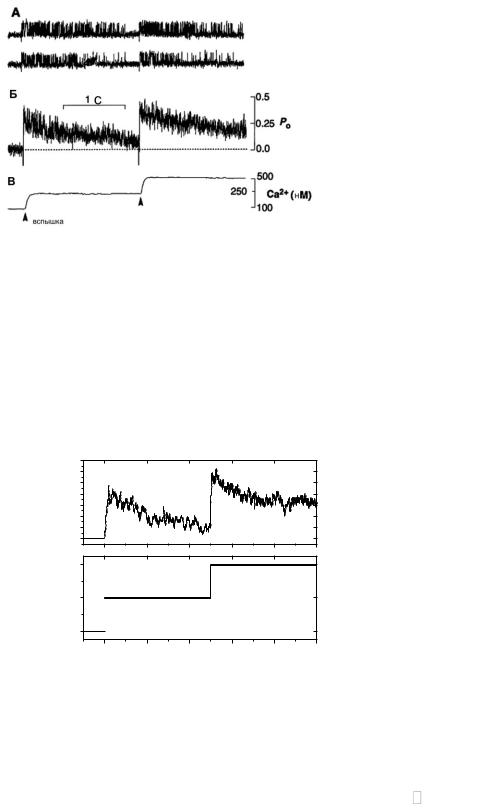
Рис. 3.25. Экспериментальное исследование влияния двух последовательных увеличений уровня cis[Ca] на активность одиночного RyR-канала. а. Реализации кальциевого тока через RyR-канал. Верхний уровень соответствует открытому состоянию канала. б. Временная зависимость вероятности пребывания канала в открытом состоянии. в. Уровень концентрации Са2+ в cis-части. Адаптировано из работы [25].
При последующих резких повышениях уровня cis[Ca] в экспериментальных работах также наблюдался эффект адаптации (рис.3.25б).
Серия численных экспериментов, проведенных в данной работе на основе ЭК-теории показала, что данная модель позволяет воспроизвести вышеописанные экспериментальные данные (рис. 3.26).
а.
|
0.7 |
|
0.6 |
|
0.5 |
open |
0.4 |
0.3 |
|
P |
0.2 |
|
|
|
0.1 |
|
0.0 |
б. |
|
0.5 |
|
|
|
|
мкМ |
|
|
cis[Ca], |
0.25 |
|
|
|
|
|
0 |
Рис. 3.26. Адаптация RyR-канала при повторной стимуляции. Временные зависимости вероятности нахождения канала в открытом состоянии (а) и уровня cis[Ca] (б). Интенсивности электронных переходов при
0<t<2.5 c:
|
|
|
|
|
|
|
|
a b |
0.01 мс 1 , |
a u |
0.01 мс-1, |
|||
|
|
|
|
|
|
|
||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|||
|
|
|
|
|
|
|
|
i b |
0.002 мс 1 , |
|
|
|
|
|
0 |
1 |
2 |
3 |
4 |
5 |
|
i u |
0.0002 мс 1 при t>2.5 c: |
||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
t, c |
|
|
|
a b |
0.025 мс 1 , |
a u |
0.01 мс-1, |
|||
|
|
|
|
|
|
|
|
|
|
|
|
|||
|
|
|
|
|
|
|
|
i b |
0.0025 мс 1 , |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
i u |
0.0002 мс 1 . |
tun |
0.002 мс 1 . |
|||
|
|
|
|
|
|
|
|
|
|
|
|
|||
Видно (рис. 3.26), что при повторном повышении cis[Ca] наблюдается медленное понижение Popen до стационарного уровня ( 0.1), при этом следует
107

указать на тот факт, что эффект адаптации проявляется даже в том случае,
когда уровень cis[Ca] перед резким повышением принимает достаточно большие значения ( 0.25 мкМ).
Вышеописанный эффект имеет большое значение для сердечной клетки, так как RyR-канал адаптируется к установившемуся уровню cis[Ca], сохраняя свою способность реагировать на следующее повышение cis[Ca]. Процесс КВВК должен быть самовосстанавливающимся, так как Са2+, высвобожденный из СР должен быть возвращен в СР и активировать следующее высвобождение Са2+,
при этом адаптация является своеобразным эффектом негативного контроля
(проверяет отсутствие нежелательных явлений и/или их повторения).
Как показывает большинство экспериментов в сердечных клетках [32, 120],
на завершение процесса высвобождения Са2+ в диадное пространство в большей степени влияет опустошение СР, но по мнению некоторых исследователей [52], понижение активности RyR-каналов во времени вследствие адаптации также вносит вклад в завершение высвобождения.
Введение впервые в данной работе в электронно-конформационную модель высокоэнергетического адсорбционного инактивационного состояния позволило описать процессы связывания ионов Са2+ с инактивационным центром RyR-канала, которые имеют меньшую вероятность, чем связывание с активационным центром. Возможность перехода в инактивацинное состояние объясняет существование длительных интервалов, наблюдаемых в эксперименте, на которых проводимость RyR-канала равна нулю [18].
3.7 Динамика одиночного RyR-канала при установившемся уровне cis[Ca]
На следующем этапе данной работы исследовалось поведение изолированных RyR-каналов при установившемся уровне cis[Ca] после выхода в динамическое равновесие (участок III на рисунке 3.23а).
Динамическое равновесие определяется постоянным значением средней вероятности пребывания RyR-канала в открытом состоянии на больших интервалах времени. Как показывают результаты экспериментов по изучению
108
кинетических свойств RyR-канала при стационарных условиях, средняя вероятность пребывания канала в открытом состоянии напрямую зависит от уровня cis[Ca] [13, 17, 20].
В связи с этим в настоящей работе необходимо было исследовать активность одиночного RyR-канала при стационарных условиях и установить способность электронно-конформационной теории объяснить эффекты динамики RyR-
канала при постоянном уровне cis[Ca], ранее выявленные экспериментально.
3.7.1 Зависимость активности RyR-канала от времени
В начале эксперимента по исследованию активности RyR-каналов все каналы в ансамбле были закрыты, а в момент времени t=0 при cis[Ca]>0
происходил быстрый процесс активации и дальнейшей процесс медленной адаптации каналов. Регистрация активности канала в численных экспериментах проводилась, после выхода на стационарный режим, начиная с момента t=3 c.
На рисунке 3.26 представлены результаты численных экспериментов по наблюдению активности RyR-канала при стационарных условиях. Данный эксперимент проводился при следующих значениях интенсивностей переходов:
|
a b |
0.1 мс-1, |
|
a u |
0.1 мс-1, |
|
i b |
0.01 |
мс-1, |
|
i u |
0.05 |
мс-1, |
|
tun |
0.05 |
мс-1. |
|
|
|
|
|
|
|
|
|
|
|
|
|
Эти значения соответствуют значению cis[Ca]=0.5 мкМ согласно формуле (2.6).
Активность канала исследовалась на малых интервалах длительностью 10 мс,
что позволило построить график зависимости вероятности пребывания RyR-
канала в открытом состоянии от времени (рис. 3.27б).
109
а. |
|
open |
|
|
б. |
|
closed |
|
|
|
0.050 |
|
|
|
|
|
|
|
|
|
open |
0.025 |
|
|
|
|
|
|
|
|
P |
|
|
|
в. |
|
0.000 |
|
|
|
|
|
|
|
|
|
H |
|
|
|
|
L |
|
|
|
|
I |
|
|
|
|
I |
|
|
|
|
3 |
t, с |
5 |
|
|
4 |
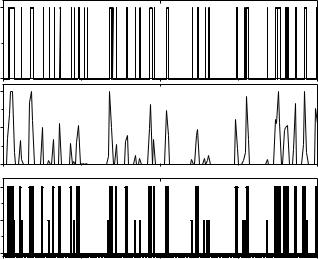
Рис. 3.27. а. Зависимость активности RyR-канала при стационарных условиях от времени. Нулевому уровню соответствует закрытое состояние. б. Зависимость вероятности пребывания канала в открытом состоянии от времени (вычислялась на временном шаге 10 мс). в. Представление активности канала в различных модах (H, L и I) от времени.
Как видно из графика, зависимость вероятности пребывания канала в
открытом состоянии P (t) является неоднородной, и динамика RyR-канала
open
при стационарных условиях носит стохастический характер.
Гистограмма распределения вероятности пребывания канала в открытом состоянии (рис. 3.28) имеет два локальных максимума в точках,
соответствующих P 0.0025 |
и |
P |
0.025. |
Локальный |
минимум |
|
open |
|
|
open |
|
|
|
соответствует значению |
P 0.006 |
и |
определяет |
граничное |
значение |
|
|
open |
|
|
|
|
|
вероятности пребывания |
канала |
в открытом состоянии Pm [121], |
которое |
|||
разделяет так называемые моды активности RyR-канала на текущем
интервале. |
При значении |
Popen Pm |
интервал |
характеризуется высокой |
|
активностью |
(мода Н), при |
0 P |
P |
канал |
находится в моде низкой |
|
|
open |
m |
|
|
активности L. Значение вероятности Popen 0 определяет нулевую активность
канала (мода I). Зависимость положения канала в модах H, L и I в зависимости от времени изображена на рисунке 3.27в. Информация о положении канала в той или иной моде позволяет более детально описать его активность в текущий момент времени.
110