

Глава 1. Клетка и ткани
 Подробное
описание этого процесса можно найти
в многочисленных руководствах по
генетике.
Подробное
описание этого процесса можно найти
в многочисленных руководствах по
генетике.
В конце интерфазы клетка практически подготовлена к митотическому делению, которое в последующем и наступает. Морфологически различают 4 фазы митоза: профаза, метафаза, анафаза и телофаза (рис. 1.1.32, 1.1.33).
Профаза характеризуется тем, что в этот период происходят интенсивные изменения структуры ядра. На участках эухроматина прекращается транскрипция. Они покрываются белками и становятся неотличимыми от зерен гетерохроматина. Затем наступает спирализа-ция хромосом. При этом хромосома становится видимой в световом микроскопе. Вышеуказанный процесс сопровождается исчезновением ядрышка. Таким образом, в начале профазы в ядре образуется плотный клубок, который к концу фазы разрыхляется, и становятся видимыми хромосомы.
Именно в этой фазе центриоли расходятся к противоположным полюсам клетки, формируется веретено, состоящее из микротрубочек.
Метафаза (следует за профазой). В мета-фазе основные изменения происходят в цитоплазме клетки. Лизосомальные ферменты растворяют ядерную оболочку, и спирализованные хромосомы оказываются в цитоплазме. Комплекс Гольджи и эндоплазматический ретикулум распадаются на мелкие фрагменты.
На каждом центромере выявляется скопление специальных белков — кинетохор. Сборка микротрубочек на материнских центриолях продолжается. В результате этого процесса формируется биполярное митотическое веретено, состоящее из микротрубочек и ассоциированных с ними белков. Различают несколько видов микротрубочек. Часть микротрубочек расходится от центриоли во все стороны. Часть их образует астральную лучистость. Другая их часть направлена к экватору клетки — полярные микротрубочки. Кроме астральных и полярных микротрубочек от полюсов отходят ки-нетохорные микротрубочки, т. е. те, которые в области экватора прикрепляются к кинетохо-рам хромосом. В клетках человека каждый кинетохор связан с 20—40 микротрубочками.
Этап формирования веретена обозначают как прометафазу. В ходе собственно метафазы хромосомы перемещаются и располагаются в одной плоскости перпендикулярно к оси между полюсами. Образуется фигура, называемая материнской звездой. В результате упорядочения положения хромосом система микротрубочек также упорядочивается. Они теперь образуют веретено деления (митотическое веретено).
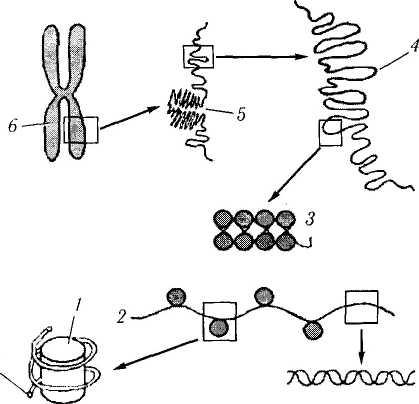
Важно отметить, что именно в метафазе определяют кариотип. У человека в норме ка-риотип характеризуется наличием 23 пар хромосом, приведенных на рис. 1.1.34. В метафазе изучают кариотип с диагностической целью при
•1 mm
U lift
90 ну
ДНК
 fl/7
П
fl/7
П
|
й |
in |
2 |
К II |
и |
|
|
ii 5& |
|
X |
6 |
|
7 8 |
9 |
|
10 |
И 12 |
|
|
пл |
Аи |
Л6 |
|
га |
АЛ |
|
|
|
13 |
14 |
15 |
16 |
17 |
18 |
19 20 |
|
л |
А А |
|
|
|
|
|
|
|
Y |
21 |
22 |
|
|
|
|
|
ДНК
Рис. 1.1.34. Кариотип человека и уровни упаковки хроматина в ядре:
а — классификация пар хромосом человека; б — молекулярная организация хромосом (по В. Л. Быкову. 1999) двойная спираль ДНК образует нить диаметром 2 нм, которая намотана на блоки дисковндшш фирмы — нуклеосомы (/), входящие в состав нук-леосомнои пиiи (2) диаметром 11 нм Скрученная нуклеосомнля нить образует хроматиновую фибриллу (3) диаметром 30 нм, которая формирует петельные домены (4) диаметром 300 нм. Более и.ютно упакованные петельные до.мены образуют конденсированные участки хромосомы (5) диаметром 700 нм, являющими ч.н тью метафазной хромосомы (б) размером около 1400 нм
различных врожденных и наследуемых заболеваниях.
Анафаза. Анафаза довольно сложный в химическом отношении процесс. В начале анафазы наступает внезапное разделение центромеры d-хромосомы, в результате чего сестринские хроматиды становятся самостоятельными s-xpo-мосомами.
Микротрубочки начинают укорачиваться, в результате чего хроматиды подтягиваются к центриолям. Сами центриоли удаляются друг от друга в сторону полюсов клетки, в результате чего образуются две дочерние звезды. В конце анафазы плазматическая мембрана как бы инвагинируется перпендикулярно к продольной оси митотического веретена, образуя борозду. В этой области под плазмолеммой появляется сократительное кольцо, состоящее из
Клетка
23

a U

Телофаза. Как указано выше, к концу анафазы и началу телофазы в середине клетки образуется цитоплазматическая перетяжка, которая постепенно углубляется и, в конце концов, полностью разделяет клетку на две равные части, содержащие идентичный набор хромосом. После этого вновь появляется ядро, а хромосомы «распадаются» с образованием глыбок хроматина. Примерно в середине телофазы начинается образование нитчатой, а затем гранулярной частей нуклеонеммы. К концу телофазы полностью сформировано ядрышко. Из мембранных пузырьков происходит формирование аппарата Гольджи, эндоплазматической сети. На этом митотический цикл завершается, и клетка входит в интерфазу.
Вышеприведенные данные характеризуют основной тип клеточного деления — митоз. Но существуют и другие типы деления. Это эндомитоз. Морфологически при эндомитозе удвоение числа хромосом происходит внутри ядерной оболочки без ее разрушения и формирования веретена деления. При повторных эндомитозах число хромосом в ядре может значительно увеличиваться и развивается так называемая полиплоидия. Сопровождается этот процесс значительным увеличением объема ядра. Полиплоидия сопровождается значительным увеличением функциональной активности клеток. Полиплоидия характерна как в норме, так и при различных патологических состояниях эндотелия роговой оболочки.
Полиплоидия развивается и при митоти-ческом делении, при котором не происходит цитотомии. При последующем делении такой двуядерной клетки хромосомные наборы ядер объединяются в метафазе, приводя к образованию двух дочерних полиплоидных клеток. Наличие полиплоидных клеток (тетра-, окта- и т.д.) является нормальным состоянием ряда тканей организма человека.
Большое биологическое значение имеет еще один тип деления — мейоз, в результате которого формируются половые клетки. Основной смысл мейоза сводится к делению, при котором достигается уменьшение количества хромосом в клетке в два раза. Обсуждение этого типа деления выходит за рамки данной книги. Более подробные сведения можно получить в большом количестве руководств по цитологии.
В литературе описан еще один тип деления — амитоз. До сих пор обсуждаются вопросы возможности существования подобного типа деления. Считают, что при таком делении исчезает биологический смысл деления, т. е. возможность равного распределения генетического материала в двух вновь образованных клетках. Тем не менее морфологи, особенно патологи, довольно часто наблюдают прямое (амитотичес-кое) деление.
1.1.5. Межклеточные соединения
Межклеточное пространство. Между цито-плазматическими мембранами соседних клеток обнаруживается равномерное светлое пространство шириной 15 нм. Это пространство нередко расширяется или сужается как в норме, так и при патологических состояниях. Несмотря на наличие межклеточного пространства, клетки довольно сильно сцеплены между собой при помощи специализированных органоидов различного типа (рис. 1.1.35). На особенностях строения этих органоидов мы и остановимся ниже.
Рис. 1 1.35. Cxi'Mii гическое изображение межклеточных контактов различного типа (по tiogan et al., 1972):
ii.iuiiii.h- кл-динеиие; 2 — :ич мосома; 3—ще.к-вой контакт I 'м .-I viz)); б — запмр.иощая зона [cmpt'-thu)
Десмосома (macula adhearens). Рядом расположенные клетки могут соединяться между собой при помощи локальных уплотнений — десмисим (рис. 1.1.36). Этот тип органоидов относится к адгезивным (контакты типа пятна слипания). При формировании подобного типа контакта цитоплазматические мембраны соседних клеток не сливаются, а как бы «прилипают» благодаря наличию межклеточного вещества. Особенно выражен подобный тип межклеточных контактов в эпителиальных тканях (эпидермис, эпителий роговицы, нейроэпите-лиальные структуры).
24