

цитология гистология и эмбриология александровская козлов радостина
.pdfА - аутосомы; Z, Y - половые хромосомы. В период размножения показаны три поколения сперматогониев и огониев, делящиеся митотически и остающиеся диплоидными.
них перестают делиться, начинают увеличиваться в размере и переходят во второй период своего развития - период роста. Другая часть спермиев продолжает делиться. Они являются стволовыми клетками и пополняют запас сперматогоний.
Период роста. В этот период клетки называются п е р в и ч н ы м и с п е р м а т о ц и т а м и . У них сохраняется диплоидное числа хромосом. В процессе роста увеличивается размер клетки и происходят сложные изменения в перераспределении наследственного материала в ядре, в связи с чем различают четыре стадии: лептотенную, зиготенную, пахитенную, диплотенную.
Для л е п т о т е н н о й (leptos - тонкий, taenia - лента) стадии характерны хорошо выраженное ядрышко, ядерная тека и спирализация хромосом, котоые имеют вид тонких нитей. Гомологичные хромосомы (отцовские и материнские) друг с другом не конъюгируют. При з и г о т е н н о й (zygoo - соединять), или синаптической (synapsis - соединение), стадии гомологичные хромосомы сближаются. Присоединившись друг к другу (конъюгировав)
51
и образовав пары - диады, обмениваются генами - кроссинговер (crossing - скрещивание, over - через). При п а х и т е н н о й (pachys - толстый) стадии пары гомологичных хромосом перекручиваются друг с другом, сильно спирализуются, поэтому укорачиваются и утолщаются. Д и п л о т е н н а я (diploos - двойной) стадия - процесс формирования тетрад. В каждой паре возникает продольная щель, и в двух конъюгировавших хромосомах образуются четыре хроматиды, которые начинают постепенно разъединяться, сохраняя на некоторое время связь друг с другом в центре своими концами. Затем эти связи теряются.
Период созревания. Это процесс развития сперматид с половинным числом хромосом, что достигается путем двух, следующих друг за другом делений созревания. После первого деления первичного сперматоцита образуются два вторичных сперматоцита, каждый из которых получает только половину от количества хромосом первичного сперматоцита. Вскоре за первым делением созревания без интерфазы (следовательно, без синтеза ДНК, характерного для периода S, в ходе которого увеличивается число хромосом) следует второе деление созревания. Из каждого вторичного сперматоцита образуется по две сперматиды. При этом пары хроматид, соединенные между собой в центре центромерой, разъединяются и распределяются по разным сперматидам, становясь хромосомами.
Следовательно, в процессе созревания из каждого первичного сперматоцита возникает четыре сперматиды с одинарным числом хромосом. Половые X- и У-хромосомы, характерные для спермиев, расходятся по разным сперматидам, в связи с чем Х- хромосомами снабжаются две сперматиды, другие две - У-хромосомами. Этот процесс обусловливает гетерогаметность самцов млекопитающих.
Способ деления, при котором дочерние клетки получают вдвое меньше число хромосом, носит название м е й о з (meiosis - уменьшение), или редукционное (reducere - уменьшение) деление.
Сперматиды имеют округлую форму, небольшие размеры, относительно крупное ядро, бедное хроматином. Митохондрии, комплекс Гольджи, центросома хорошо развиты и
расположены вблизи ядра. Другие органеллы, а также включения почти отсутствуют. Сперматиды не способны делиться.
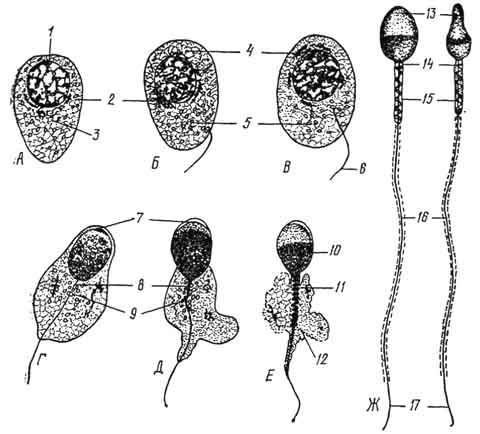
Период формирования (рис. 39). Завершающий период сперматогенеза, в ходе которого округлая клетка - сперматида - приобретает морфологические свойства, характерные для спермия. В ходе формирования светлое округлое ядро сперматиды постепенно уплотняется, становится овальным и перемещается к плазмолемме. Комплекс Гольджи передвигается к ядру и продуцирует уплотеннную гранулу - а к р о б л а с т . Последний увеличивается в размере и в виде чехлика охватывает ядро сперматиды - будущую головку спермия.
В средней зоне акробласта из мельчайших зерен формируется а к р о с о м а . Это уплотненное тельце, богатое ферментом гиалуронидазой. К противоположному от ядра полюсу перемещается центросома, в которой различают проксимальную и дистальную
52
Рис. 39. Период формирования (схема). Дифференцировка сперматиды и образование спермия (А -
Ж).
1 - ядро сперматиды; 2 - комплекс Гольджи; 3 - центриоли; 4 - зачаток акросомы; 5 - митохондрии; 6 - жгутик; 7 - акросомный чехлик; 8 - проксимальная центриоль; 9 - дистальная центриоль; 10 - ядерное вещество, сконцентрированное в головке спермия; 11 - митохондриальная спираль; 12 - остатки цитоплазмы; 13 - головка; 14 - шейка; 15 - средняя часть; 15 - хвостик; 17 - концевой участок хвостика.
ц е н т р и о л и . Они образуют шейку спермия. Дистальная центриоль делится на две части - переднюю и заднюю. От передней части дистальной центриоли отрастает осевая нить хвостика, состоящая из микротрубочек. Задняя часть дистальной центриоли приобретает вид колечка. Она, сползая по осевой нити хвостика и увлекая за собой цитоплазму сперматиды, содержащую митохондрии и гликоген, располагается на границе между
начальным и главным отделами хвостика спермия. Смещаясь по хвостику, цитоплазма тонким слоем одевает главную его часть. Клетка продолжает удлиняться и приобретает форму спермия.
Развитие яйцеклеток, или овогенез (см. рис. 38 - II), - сложный и очень длительный процесс. Он начинается в период эмбриогенеза и завершается в органах половой системы половозрелой самки. Слагается овогенез из трех периодов: размножения, роста, созревания.
Период размножения протекает в период внутриутробного развития и завершается в течение первых месяцев после рождения. Клетки называются овогониями (оогониями). Они
53
незначительных размеров, бедны цитоплазмой, их ядра имеют диплоидное число хромосом. Оогонии интенсивно митотически делятся. Некоторые из них перестают делиться, покрываются клетками фолликулярного эпителия, которые снабжают яйцеклетку питательным материалом.
В период роста клетки называются п е р в и ч н ы м и о о ц и т а м и . Они увеличиваются в размере. Изменения в ядрах аналогичны первичным сперматоцитам; они проходят также лептотенную, зиготенную, пахитенную и диплотенную стадии. Затем в ооците начинается интенсивный синтез и накопление желтка, необходимого для течения первых стадий развития зародыша, в связи с чем хромосомы вновь деспирализуются. Этот процесс протекает в две фазы: стадия малого роста (превителлогенез) и стадия большого роста (вителлогенез). Морфологические особенности п р е в и т е л л о г е н е з а - локализация органелл вокруг ядра, активизация функции белоксинтезирующей системы цитоплазмы: развиваются рибосомы, гранулярная эндоплазматическая сеть, поэтому цитоплазма окрашивается основными красителями, то есть она базофильна. Хорошо развиты комплекс Гольджи и митохондрии. Ядро округлое, содержит мало хроматина, хорошо развито ядрышко. Вторичная оболочка ооцита состоит из одного слоя фолликулярных клеток. Превителлогенез обычно длится до наступления половой зрелости самки.
В и т е л л о г е н е з - стадия интенсивного накопления желтка в цитоплазме первичного ооцита. В данном процессе принимает участие весь организм. Фолликулярные клетки в ооцит транспортируют желток. Они интенсивно делятся митозом, поэтому вторичная оболочка становится многослойной.
Органеллы из перинуклеарной зоны перемещаются на периферию: формируется кортикальный слой, который играет важную роль в начальных стадиях эмбриогенеза.
Период созревания состоит из двух быстро следующих друг за другом делений созревания, в ходе которых диплоидная клетка становится гаплоидной. Этот процесс обычно протекает в яйцеводе после выхода первичного ооцита из яичника.
Первое деление созревания завершается образованием двух неравноценных структур - вторичного ооцита и первого направительного, или редукционного, тельца. Каждая из них по сравнению с первичным ооцитом содержит половину количества ядерного вещества. За первым делением очень скоро наступает второе, в ходе которого также образуются только одна зрелая яйцеклетка и второе неправильное тельце. Первое направительное тельце к этому моменту делится еще один раз и образуются два направительных тельца.

Следовательно, из одного первичного ооцита в процессе созревания возникают только одна зрелая яйцеклетка и три направительных тельца.
Таким образом, овогенез отличается от сперматогенеза рядом особенностей: начинается в период внутриутробного развития
54
самки, имеет длительный период роста, в процессе которого накапливается питательный материал (желток); протекает в яичнике эмбриона и половозрелого животного, а затем в яйцеводе; из одного первичного ооцита образуется только одна зрелая яйцеклетка; все яйцеклетки генетически однородны, так как имеют Х-хромосому; женская половая клетка не способна к самостоятельному передвижению.
55
50 :: 51 :: 52 :: 53 :: 54 :: 55 :: Содержание

55 :: 56 :: 57 :: 58 :: Содержание
РАННИЕ ПЕРИОДЫ ЭМБРИОНАЛЬНОГО РАЗВИТИЯ (ЭМБРИОГЕНЕЗ)
ОПЛОДОТВОРЕНИЕ
Оплодотворение - слияние половых гамет (спермия и яйцеклетки) и образование нового одноклеточного организма (зиготы) или оплодотворенной яйцеклетки (рис. 40). От зрелой яйцеклетки она отличается удвоенной массой ДНК, диплоидным числом хромосом, высокой метаболической активностью. У многих животных оплодотворение яйцеклетки внутреннее, и оно протекает в яйцеводе при пассивном ее продвижении по направлению матки. Движение спермиев в половых путях самки осуществляется благодаря функции аппарата движения этой клетки, положительного хемотаксиса (движение по направлению химических раздражителей), реотаксиса (движение против тока жидкости), перистальтическим сокращениям стенки матки; движению ресничек клеток, покрывающих внутреннюю поверхность яйцевода.
При сближении половых клеток ферменты акросомы головки спермия разрушают вторичную оболочку яйцеклетки. В момент прикосновения спермия к плазмолемме яйцеклетки на ее поверхности образуется выпячивание цитоплазмы - воспринимающий бугорок, или бугорок оплодотворения. В ооцит проникают головка, шейка, несущая центросому, и начальная часть хвостового отдела. У млекопитающих в оплодотворении участвует только один спермий. Этот процесс носит название м о н о с п е р м и и (рис. 41). Как уже говорилось выше, яйцеклетки всегда содержат только Х-половую хромосому, а спермии либо Х-половую хромосому, либо Y-половую хромосому. Если в яйцеклетку проникает спермий с Y-половой хромосомой, то зигота будет содержать Х,У-половые хромосомы, и развивается самец. При наличии в зиготе Х,Х-половых хромосом будет развиваться самка.
Рис. 40. Оплодотворение яйцеклетки спермием (схема).
55
У птиц, рептилий, хвостатых амфибий наблюдается п о л и с п е р м и я : в яйцеклетку проникает одновременно несколько спермиев. У птиц все спермии имеют Z-хромосому, а яйцеклетки или Z-половую хромосому, или W-хромосому. Таким образом, новый организм получает хромосомы мужской и женской половых клеток и поэтому наследует признаки обоих родителей.
После проникновения спермия в яйцеклетку вокруг последней формируется оболочка оплодотворения, препятствующая проникновению в ооцит других спермий. Яйцеклетки выделяют особые вещества - агглютинины. Они склеивают другие спермии, а фолликулярные клетки вторичной оболочки яйцеклетки их поглощают.
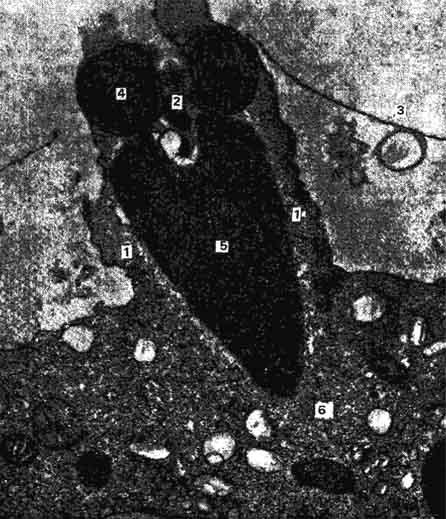
Рис. 41. Спермий, втягиваемый в цитоплазму яйцеклетки воспринимающим бугорком (электронная микрофотография) :
1 - воспринимающий бугорок; 2 - центриолъ спермип; 3 - оболочка оплодотворения; 4 - мнтохондриальное тельце; 5 - ядро спермия; 6 - цитоплазма яйцеклетки (по Вилли и Детье).
56
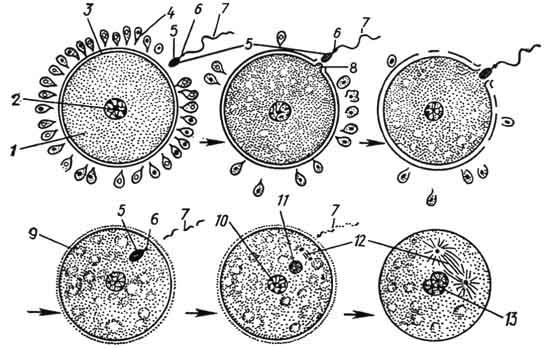
Рис. 42. Схематическое изображение последовательных стадий оплодотворения:
1 - цитоплазма яйцеклетки; 2 - ядро яйцеклетки; 3 - блестящая оболочка; 4 - фолликулярный эпителий; 5 - головка спермия; 6 - шейка спермия; 7 - хвост спермия; 8 - воспринимающий бугорок; 9 - оболочка оплодотворения; 10 - женский и 11 - мужской пронуклеусы; 12 - веретено между центриолями; 13 - синкарион (по Елисееву).
Головка спермия, внедрившаяся в цитоплазму яйцеклетки, набухает, округляется и приобретает форму округлого ядра - мужской пронуклеус (рис. 42). Последний перемещается к округлому ядру яйцеклетки - женскому пронуклеусу. Оба пронуклеуса гаплоидны, и после их соединения образуется диплоидное ядро зиготы. Процесс соединения пронуклеусов называется с и н к а р и о н о м . В это время в клетке резко усиливаются метаболические процессы.
Яйцеклетка лишена центриолей. В нее вносят их спермии. Центриоли шейки спермия расходятся, образуется ахроматиновое веретено; ядерная мембрана исчезает, отцовские и материнские хромосомы формируют материнскую звезду первого митотического деления оплодотворенной яйцеклетки. Так начинается следующий этап эмбриогенеза - дробление.
При полиспермии (рептилии, птицы и др.) в яйцеклетку проникает одновременно несколько спермиев, однако с ее ядром соединяется только один из них. Неоплодотворенные яйцеклетки и неучаствующие в оплодотворении спермии гибнут и в яйцеводах или матке поглощаются клетками крови - лейкоцитами (leukos - белый, kytos - клетка).
Таким образом, при оплодотворении осуществляется: 1) активация яйцеклетки, после чего начинается ее деление (дробление);
57
2) рекомбинация отцовских и материнских хромосом, что в конечном результате обусловливает адаптацию особи к изменениям окружающей среды и их выживаемость. Таковообщебиологическое значение оплодотворения.
58

55 :: 56 :: 57 :: 58 :: Содержание

58 :: 59 :: 60 :: 61 :: 62 :: Содержание
ДРОБЛЕНИЕ
Дробление - дальнейший процесс развития одноклеточной зиготы, в ходе которого образуется многоклеточная бластула, которая состоит из стенки - б л а с т о д е р м ы и полости - б л а с т о ц е л я . В бластодерме различают крышу, дно и расположенную между ними краевую зону. В процессе митотического деления зиготы образуются новые клетки - бластомеры, остающиеся тесно связанными друг с другом.
В начальной стадии дробления многоклеточный организм по своему размеру сходен с зиготой, так как его бластомеры, делясь, не достигают размера исходной клетки. Характер дробления в. эволюционном ряду хордовых различен, что в значительной степени обусловлено количеством и распределением желтка в яйцеклетках.
Дробление может быть полным (голобластическим) или частичным (меробластическим). При голобластическом дроблении принимает участие весь материал зиготы, при меробластическом - только та ее зона, которая лишена желтка.
Полное дробление классифицируют на равномерное и неравномерное. П о л н о е р а в н о м е р н о е д р о б л е н и е (рис. 43) характерно для яиц с малым количеством желтка (олиголецитальных) и равномерно расположенным по всей цитоплазме клетки желтком (изолецитальных). Примером такого дробления могут служить ланцетник, аскарида и др. В оплодотворенной яйцеклетке различают два полюса: верхний - анимальный и нижний - вегетативный.
После оплодотворения желток, незначительное количество которого было равномерно распределено то всей цитоплазме, перемещается к вегетативному полюсу. Первая борозда дробления проходит в меридиональном направлении и делит зиготу на два бластомера, которые соответствуют будущей левой и правой половине тела зародыша. Вторая борозда дробления проходит также меридионально под прямым углом к первой, и теперь зародыш состоит из четырех бластомеров. Третья борозда дробления имеет экваториальное направление, поэтому каждый бластомер делится на две части. Такой зародыш построен из восьми бластомеров, при этом четыре из них образовались из вегетативного полюса зиготы, в связи с чем они содержат весь желток зиготы и отличаются большими размерами. Эти бластомеры соответствуют задней части тела; анимальные - четыре - передней части.
Затем появляются две меридиональные борозды, делящие зародыш на 16 бластомеров. Пятое дробление - это две широтные борозды, в составе зародыша 32 бластомера. Они начинают
58
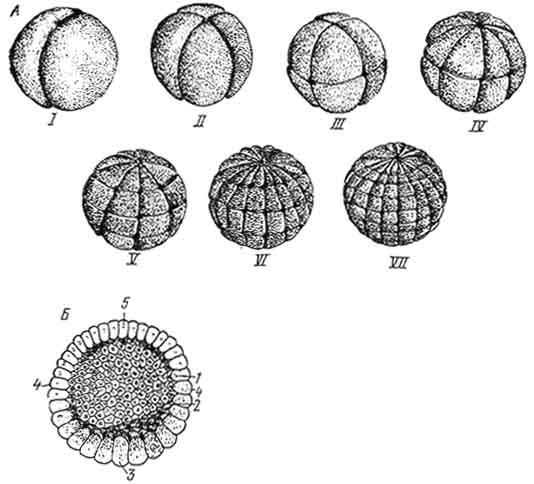
Рис. 43. Схема расположения борозд дробления у ланцетника (A):
I - зародыш на стадии двух бластомеров; II - зародыш на стадии четырех бластомеров; III - зародыш на стадии восьми бластомеров; IV - зародыш на стадии 16 бластомеров; V - зародыш на стадии 32 бластомеров; VI - зародыш на стадии 64 бластомеров; VII - зародыш на стадии 128 бластомеров. Строение бластулы (Б): 1 - бластодерма; 2 - бластоцель; 3 - дно; 4 - краевая зона; 5 - крыша бластулы.
постепенно отодвигаться друг от друга, контактируя лишь боковыми поверхностями. Внутри зародыша образуется сначала небольшая полость - бластоцель, которая постепенно увеличивается. После шестого дробления образуется 64 клетки, при этом борозды дробления проходят меридионально. После седьмого дробления (возникают четыре широтные борозды) зародыш состоит из 128 бластомеров.
Позднее синхронность в делении зародыша нарушается, бластомеры отодвигаются на периферию и располагаются в один слой, формируя бластодерму, а в центре зародыша образуется бластоцель.
Дробление завершается образованием бластулы, форма которой напоминает шар, заполненный жидкостью. Стенка шара образована клетками бластодермы.
Таким образом, при полном равномерном дроблении материал всей зиготы участвует в делении и после каждого деления (дробления) число клеток (бластомеров) увеличивается вдвое.
В бластодерме дифференцируются следующие участки: крыша, построенная из относительно мелких бластомеров; дно - это более крупные бластомеры и краевая зона, лежащая между дном и крышей бластулы.