

цитология гистология и эмбриология александровская козлов радостина
.pdfсубстанция (эухроматин) исчезает. Функциональные неактивные участки хромосом уплотнены и микроскопически выявляются как базофильные глыбки хроматина (гетерохроматин). Количество и локализация конденсированного хроматина хромосом интерфазных ядер различных типов клеток соответствует специфичности активности их белкового обмена. Конденсированный хроматин преимущественно выявляется на внутренней поверхности ядерной оболочки и в разной степени в виде глыбок в других зонах ядра. Максимально конденсируется хроматин в период митотического деления клеток, когда хромосомы выключаются из процессов синтеза, уплотняясь, формируют характерные для каждого вида клеток наборы хромосом.
17
Ядрышко - тельце сферической формы диаметром 1 - 5 мкм, сильно преломляющее свет. Размеры его варьируют в зависимости от физиологического состояния клеток. Наиболее крупные ядрышки встречаются в быстро размножающихся эмбриональных клетках и клетках опухолей. Формирование ядрышка зависит от специфического участия хромосомы - я д р ы ш к о в о г о о р г а н и з а т о р а (рис. 11). Число ядрышек в ядре соответствует числу ядрышковых организаторов. Последние обычно располагаются в области вторичных перетяжек хромосом и содержат гены, кодирующие синтез рибосомальной РНК. Ядрышко окрашивается кислыми и особенно основными красителями.
Функция ядрышек - формирование рибосом. При небольших увеличениях электронного микроскопа в ядрышке обнаруживают а м о р ф н у ю ч а с т ь и н у к л е о л о н е м у (ядрышковая нить), представляющую собой сеть нитей толщиной 60 - 80 нм. При больших увеличениях электронного микроскопа можно видеть, что аморфная часть состоит из филаментов толщиной 5 - 8 нм, а нуклеолонема построена из филаментов толщиной 5 - 8 нм и гранул диаметром 15 - 20 нм. Филаменты и гранулы состоят из РНК. На периферии ядрышка располагается околоядрышковый хроматин. Его рассматривают как конденсированную часть хроматина ядрышкового организатора. Электронномикроскопические исследования с использованием меченого уридина показали, что сначала метка включается в фибриллы, а затем в гранулы. Следовательно,

Рис. 10. Схема различных уровней строения ДАП:
1 - нуклеосомы; 2 - межнуклеосомальные участки ДНК; 3 - фибрилла ДНП с диаметром 20 - 25 нм (по Ченцову).
Рис. 11. Схема организации ядрышка:
1 - околоядрышковый хроматин; 2 - ДНК ядрышкового организатора (область ДНК с рибосомальными генами); 3 - фибриллярная зона; 4 - гранулярная зона; 5 - белки и РНП ядрышкового матрикса.
18
вначале формируются фибриллы, затем конфигурация их изменяется и они превращаются в гранулы. В ядрышке РНК связывается с белком. Здесь происходит сборка субъединиц

рибосом, которые, по-видимому, и являются гранулярным компонентом ядрышка. Окончательное формирование рибосом происходит вые ядрышка.
Ядерный сок (кариоплазма) - микроскопически бесструктурное вещество ядра. Он содержит различные белки (нуклеопротеиды, гликопротеиды), ферменты и соединения, участвующие в процессах синтеза нуклеиновых кислот, белков и других веществ, входящих в состав кариоплазмы. Электронно-микроскопически в ядерном соке выявляют рибонуклеопротеидные гранулы 15 нм в диаметре.
В ядерном соке выявлены гликолитические ферменты и их субстраты, участвующие в синтезе и расщеплении свободных нуклеотидов и их компонентов, энзимы белкового и аминокислотного обмена и др. Сложные процессы жизнедеятельности ядра обеспечиваются энергией, освобождающейся в процессе гликолиза, ферменты которого содержатся в ядерном соке.
19
14 :: 15 :: 16 :: 17 :: 18 :: 19 :: Содержание

19 :: 20 :: 21 :: 22 :: 23 :: 24 :: 25 :: 26 :: 27 :: 28 :: 29 :: 30 :: 31 :: 32 :: 33 :: 34 :: 35 :: 36 :: 37 :: 38 :: Содержание
ЦИТОПЛАЗМА
Цитоплазма клетки состоит из микроскопически бесструктурного основного вещества - гиалоплазмы, в которой рассредоточены ее специализированные структуры (органеллы), выполняющие специфические функции.
Гиалоплазма - гетерогенное по химическому составу вещество цитоплазмы клеток. Оно содержит белки, нуклеиновые кислоты, полисахариды, аминокислоты, нуклеотиды, различные ферменты и многие другие соединения, участвующие в метаболизме клеток. Гиалоплазма - среда, объединяющая различные структуры клетки и обеспечивающая их взаимодействие. В гиалоплазме сосредоточены АТФ, продукты обмена, включения глыбок гликогена, капель жира, пигменты и др.
Органеллы - структуры цитоплазмы, выполняющие в клетке специфические функции. К ним относят плазмолемму, рибосомы, эндоплазматическую сеть, комплекс Гольджи, лизосомы, пероксисомы, митохондрии, центриоли. Кроме названных органелл, в цитоплазме клеток имеется значительное количество различных по морфологии и функциональному значению структур (фибрилл, филаментов, микротрубочек), отражающих специфичность дифференцировки, характерной для определенных тканей.
Плазмолемма - оболочка клетки, выполняющая ограничительную, транспортную и рецепторную функции. Она отграничивает клетку с поверхности и обеспечивает связь с внешней средой. Плазмолемма обеспечивает механическую связь клеток и межклеточные взаимодействия, содержит клеточные рецепторы гормонов и других сигналов окружающей клетку среды, осуществляет транспорт веществ в клетку и из клетки как по градиенту
19
концентраций - пассивный перенос, так и с затратами энергии против градиента концентраций - активный перенос.
В состав оболочки входят плазматическая мембрана, надмембранный комплекс - гликокалекс и субмембранный опорно-сократительный аппарат. В основе плазматической мембраны лежит бимолекулярный слой липидов, в который погружены полностью тали частично молекулы белка и гликопротеидов. В ней около 40% липидов, 60% белков и до 1% углеводов. В связи с функциональной характеристикой клеток различных тканей специфичен состав тликопротеидного надмембранного комплекса. В нем содержится до 1% углеводов (гиалуроновая, сиаловая кислоты и др.), молекулы которых образуют длинные ветвящиеся цепи полисахаридов, связанные с белками мембраны (рис. 12 и 13). Находящиеся в гликокалексе белки - ферменты участвуют в конечном внеклеточном расщеплении веществ. Продукты этих реакций в виде мономеров поступают в клетку. При активном переносе транспорт веществ в клетку (э н д о ц и т о з ) осуществляется или поступлением молекул в виде раствора - пиноцитоз, или захватом крупных частиц - фагоцитоз.
Процесс ф а г о ц и т о з а состоит из двух фаз: взаимодействие частицы с рецептором плазмолеммы клетки и затем поглощение ее в результате образования псевдоподий. Первоначальное взаимодействие частицы и рецептора плазмолеммы вызывает сигнал, б результате которого происходят местные скопления контрактильных белков (актина и др.) в поверхностном слое цитоплазмы, ведущие к образованию псевдоподий. Это увеличивает

площадь ее контакта с частицей, что вызывает дальнейшее скопление сократимых белков. Процесс продолжается, пока псевдоподии не сомкнутся над частицей, формируя фагосому.
П и н о ц и т о з - везикулярное поглощение жидкости, содержащей низкомолекулярные растворы (липопротеиды, иммунные комплексы, ферритин, гормоны и др.). Различают макропиноцитоз, при котором ундулирующие складки поверхности клетки захватывают капли раствора, видимые при фазоконтрастной микроскопии, и микропиноцитоз - жидкость захватывается минимальными инвагинациями, различимыми только при электронной микроскопии.
По механизму действия микропиноцитоз бывает жидкофазный и абсорбтивный. Первый - неизбирательный: растворенные вещества поглощаются пропордионально их концентрации в жидкой среде, а поглощающая их мембрана морфологически не специализирована. При втором - мембраны пузырькообразпых инвагинаций плазмолеммы клетки покрыты с внешней поверхности тонким местным слоем гликокалекса, а с внутренней - щетинкой тонких волосков.
Количество иитериоризированной (погруженной) при эндоцитозе мембраны может быть большим, особенно при фагоцитозе. Макрофаги in vitro могут интериоризировать при фагоцитозе до 18% своей плазмолеммы в час.
20
Рис. 12. Схема строения плазматической мембраны: белки с внешней стороны слоя связаны с полисахаридами, образуя слой гликокалекса.
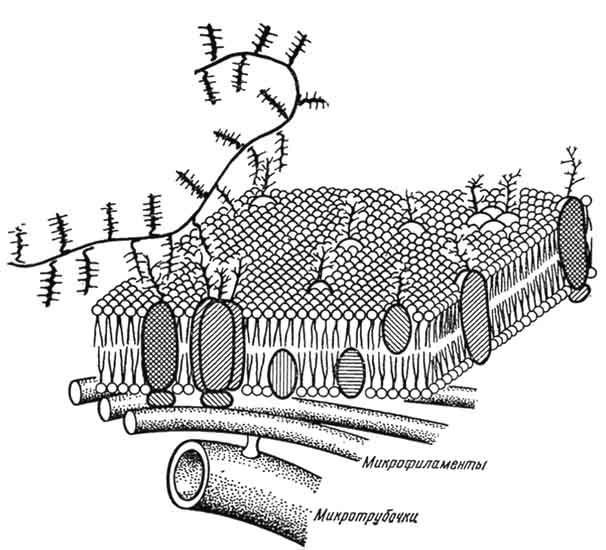
Рис. 13. Гипотетическая схема плазматической мембраны (по Бергельсону).
21

Рис. 14. Схема клеточных контактов:
1 - простой контакт; 2 - замок; 3 - плотный контакт; 4 - промежуточный контакт; 5 - десмосома; б - щелевой контакт.
В соответствии с функциональными и морфологическими особенностями тканей оболочка клеток образует характерные для них аппараты м е ж к л е т о ч н ы х к о н т а к т о в . Основные их формы: простой контакт, плотный контакт, промежуточный контакт (или зона слипания) и щелевой контакт (рис. 14).
Простой контакт - наиболее распространенная форма контакта двух смежных клеток. При нем клетки отстоят одна от другой на расстоянии 15 - 20 нм. Межклеточное пространство соответствует надмембранным компонентам клеточных мембран контактирующих клеток.
Плотный (замыкающий) контакт. При нем внешние слои плазмолеммы у люминальной поверхности смежных клеток сливаются в одну общую структуру и изолируют межклеточное пространство от внешней для ткани среды. Этот тип соединения находится между эпителиальными клетками у их апикальной поверхности и образует зону слияния мембран (слипания их интегральных белков), окружающую в виде пояска верхушки клеток. Белки мембраны связаны в зоне плотного замыкающего контакта с системой тонких фибрилл цитоплазмы, ориентированных параллельно поверхности клетки по ходу зоны слипания.
Разновидностью плотного контакта являются десмосомы. Они характеризуются особым развитием и дифференцировкой надмембранного комплекса смежных клеток. В точечных десмосомах расстояние между мембранами двух контактирующих клеток 22 - 35 нм. В межклеточном пространстве за счет надмембранного комплекса формируется
волокнистое вещество. В его центральной части образуется пластинка, содержащая белки и мукополисахариды. Она связана с плазмолеммами смежных клеток поперечными фибриллами. К мембранам контактирующих клеток прилегают электроноплотные зоны цитоплазмы с отходящими от них фибриллами. Десмосомы обеспечивают механическую связь смежных клеток.
Щелевой контакт характеризуется наличием незначительного межклеточного пространства (до 2 - 3 нм). Это специализированная область плазмолемм смежных клеток, обеспечивающая диффузию ионов и мелких молекул от одной клетки в другую. При соответствующей обработке ткани электроноплотным веществом видно, что межклеточное пространство пересекается мостиками
22
диаметром 7 нм на расстоянии до 10 нм. В некоторых случаях в мостиках отмечают наличие мельчайших пор. Соответствующие материалы получены и замораживанием - сколом. Это дает основание полагать, что глобулярная частица в области щелевого контакта тянется через бислой липидов мембран и впячивается в межклеточную щель, где соединяется с соответствующей частицей противоположной мембраны смежной клетки. Соединение конец в конец этих частиц образует единицы - коннексоны, по которым из клетки в клетку идет гидрофильный канал диаметром 1,5 - 2 нм, проводящий ионы и мелкие молекулы, поддерживая их электрические и метаболические взаимодействия. Проницаемость щелевых контактов достоверно доказывается прохождением при микроинъецировании флуоресцентных красителей, аминокислот, нуклеотидов и других веществ из одной клетки в другую. Белки, аминокислоты и другие макромолекулы через щелевой контакт не проходят.
Рибосомы представляют собой гранулы 15 - 35 нм в диаметре. Располагаются они в цитоплазме свободно или фиксированы на мембране эндоплазматической сети (гранулярная эндоплазматическая сеть). Свободные рибосомы характерны для цитоплазмы недифференцированных камбиальных клеток. При световой микроскопии цитоплазма клеток, богатых рибосомами, базофильна. Рибосомы имеются и в составе ядра, где они обеспечивают синтез ядерных белков (рис. 15).
Состоят рибосомы из двух субъединиц - малой и большой. Малая субъединица прикреплена к уплощенной области большой субъединицы. Каждая из них содержит молекулу рибосомальной РНК (р-РНК) и белка, который составляет 40 - 60% общей массы рибосомы. Располагаясь на мембранах эндоплазматической сети цитоплазмы клетки, рибосома прикрепляется большой субъединицей.
Рибосомы участвуют в сборке молекул белка - укладке аминокислот в полимерные цепи в строгом соответствии с генетической информацией, заключенной в ДНК.
Помимо рибосомальной РНК, в клетке присутствует информационная РНК (и-РНК), синтезирующаяся на ДНК ядра. Последняя определяет порядок чередования азотистых оснований в и-РНК. и-РНК несет информацию от генома к рибосомам цитоплазмы, где закодированное сообщение транслируется в последовательность включения аминокислот синтезируемого белка.
Белок синтезируется обычно не на одной рибосоме, а на группе рибосом - полирибосоме (п о л и с о м е ). Рибосомы в полисоме связаны молекулой и-РНК, которая проходит вдоль ряда рибосом, пока вся закодированная в ней информация не будет
прочитана. Информационная РНК связана с малой субъединицей рибосом, формирующаяся полипептидная цепочка - с большой.
Третий вид РНК в цитоплазме - это транспортная РНК (т-РНК), которая переносит аминокислоты на рибосому. Существует специальная т-РНК для каждой аминокислоты, и каждая
23

Рис. 15. Полирибосомы ретикулоцитов: А - напыленные платиной (ув. 100000); Б - окрашенные позитивно уронилацетатом (ув. 400000, по Рпч)
24