

цитология гистология и эмбриология александровская козлов радостина
.pdfРис. 88. А - клетка Лангерганса; Б - специфические гранулы "теннисные ракетки с ампулярным концевым расширением и продольными ламеллами в области рукоятки". Электронная микрофотография.
З е р н и с т ы й с л о й (см. рис. 79, е - в) состоит из 2 - 4 рядов клеток плоской формы, лежащих параллельно поверхности эпителиального пласта. Для эпителиоцитов характерны округлые, овальные или вытянутые ядра; уменьшение количества органелл; накопление кератиногиалинового вещества, пропитывающего тонофибриллы. Кератогиалин окрашивается основными красителями, поэтому имеет вид базофильных гранул. Кератиноциты
Рис. 89. Клеточные мостики в эпидермисе носового зеркальца быка:
1 - ядро; 2 - клеточные мостики.
113
"зернистого слоя являются предшественннками клеток следующего - блестящего слоя (е - г). Его клетки лишены ядер и органелл, а тонофибриллярно-кератиногиалиновые комплексы сливаются в гомогенную массу, сильно преломляющую свет и
окрашивающуюся кислыми красителями. Электронно-микроскопически этот слой не выявлен, так как не имеет ультраструктурных отличий.
Р о г о в о й с л о й (е - д) состоит из роговых чешуек. Они образуются из блестящего слоя и построены из кератиновых фибрилл и аморфного электроноплотного материала, роговой слой снаружи покрыт однослойной мембраной. В поверхностных зонах фибриллы лежат более плотно. Роговые чешуйки соединяются .друг с другом с помощью ороговевших десмосом и других структур клеточных контактов. Потеря роговых чешуек возмещается новообразованием клеток базального слоя.
Итак, кератиноциты поверхностного слоя превращаются в плотное неживое вещество - кератин (KERATOS - рог). Он защищает нижележащие живые клетки от сильных механических воздействий и высыхания. Кератин препятствует вытеканию тканевой жидкости из межклеточных щелей.
Роговой слой выполняет функцию первичного защитного барьера, так как он непроницаем для микроорганизмов. Ороговевающий плоский и многослойный эпителий может достигать значительной толщины, что приводит к нарушению питания его клеток. 'Это устраняется образованием соединительнотканных выростов - сосочков, которые увеличивают поверхность контакта клеток базального слоя и рыхлой соединительной ткани, выполняющей трофическую функцию.
Переходный эпителий (ж) развивается из мезодермы и подрывает внутреннюю поверхность почечной лоханки, мочеточников, мочевого пузыря. При функционировании этих органов меняется - объем их полостей, в связи с чем толщина эпителиального пласта то резко снижается, то возрастает.
Эпителиальный пласт состоит из базального, промежуточного, поверхностного слоев
(ж - а, б, в).
Базальный слой построен из базальных клеток, связанных с базальной мембраной, различных по форме и размеру: мелкие -кубические и крупные грушевидные клетки. Первые из них имеют округлые ядра и базофильную цитоплазму. В эпителиальном пласте ядра этих клеток образуют самый нижний ряд ядер. Мелкие кубические клетки характеризуются высокой митотической активностью и выполняют функцию стволовых клеток. Вторые - своей узкой частью прикреплены к базальной мембране. Их расширенное тело расположено над кубическими клетками; цитоплазма светлая, так как слабо выражена базофилия. Если орган не наполнен мочой, крупные грушевидные клетки нагромождаются друг на друга, формируя как бы промежуточный слой.
Покровные клетки уплощены. Часто многоядерны или их ядра полиплоидны (содержат большее количество хромосом по
114

Рис. 90. Переходный эпителий почечной лоханки овцы:
а - а' - слизистая клетка покровной зоны со слабой реакцией на слизь; б - промежуточная зона; в - митоз; г - базальная зона: д - соединительная ткань.
Рис. 91. Переходный эпителий мочевого пузыря кролика:
1 - в спавшемся; 2 - в слаборастянутом; 3 - в сильнорастянутом мочевом пузыре.
115

сравнению с диплоидным набором хромосом). Поверхностные клетки могут ослизняться. Эта способность особенно хорошо развита у травоядных (рис. 90). Слизь предохраняет эпителиоциты от вредных воздействий мочи.
Таким образом в перестройке эпителиального пласта данного вида эпителия играет степень наполнения органа мочой (рис.91).
116
103 :: 104 :: 105 :: 106 :: 107 :: 108 :: 109 :: 110 :: 111 :: 112 :: 113 :: 114 :: 115 :: 116 :: Содержание

116 :: 117 :: 118 :: 119 :: Содержание
ЖЕЛЕЗИСТЫЙ ЭПИТЕЛИЙ
Способность клеток организма интенсивно синтезировать активные вещества (секрет, гормон), необходимые для осуществления функции других органов, характерна для эпителиальной ткани. Эпителии, вырабатывающие секреты, называются железистыми, а его клетки - секреторными клетками, или секреторными гландулоцитами. Из секреторных клеток построены железы, которые могут быть оформлены в виде самостоятельного органа или являться только его частью.
Различают эндокринные (ENDO - внутри, KRIO - отделяю) и экзокринные (ехо - снаружи) железы. Э к з о к р и н н ы е железы состоят из двух частей: концевой (секретирующей) части и выводных протоков, по которым секрет поступает на поверхность организма или в полость внутреннего органа. Выводные протоки обычно не принимают участие в образовании секрета.
Э н д о к р и н н ы е железы лишены выводных протоков. Их активные вещества (гормоны) поступают в кровь, в связи с чем функцию выводных протоков выполняют капилляры, с которыми железистые клетки очень тесно связаны. Подробно функциональная морфология желез внутренней секреции будет рассмотрена в главе 8.
Экзокринные железы разнообразны по строению и функции. Они могут быть одноклеточными и многоклеточными. Примером одноклеточных желез служат бокаловидные клетки, встречающиеся в простом столбчатом каемчатом и псевдомногослойном реснитчатом эпителиях. Несекретирующая бокаловидная клетка цилиндрической формы и сходна с несекреторными эпителиоцитами. Секрет (муцин) накапливается в апикальной зоне, а ядро и органеллы смещаются к базальной части клетки. Смещенное ядро приобретает форму полулуния, а клетка - бокала. Затем секрет изливается из клетки, а она вновь приобретает столбчатую форму.
Экзокринные многоклеточные железы могут быть однослойными и многослойными, что обусловлено генетически. Если железа развивается из многослойного эпителия (потовая, сальная, молочная, слюнные железы), то и железа многослойна; если из однослойного (железы дна желудка, матки, поджелудочная железа), то они однослойны.
Характер ветвления выводных протоков экзокринных желез различен, поэтому они подразделяются на простые и сложные. У простых желез неветвящийся выводной проток, у сложных - ветвящийся.
116
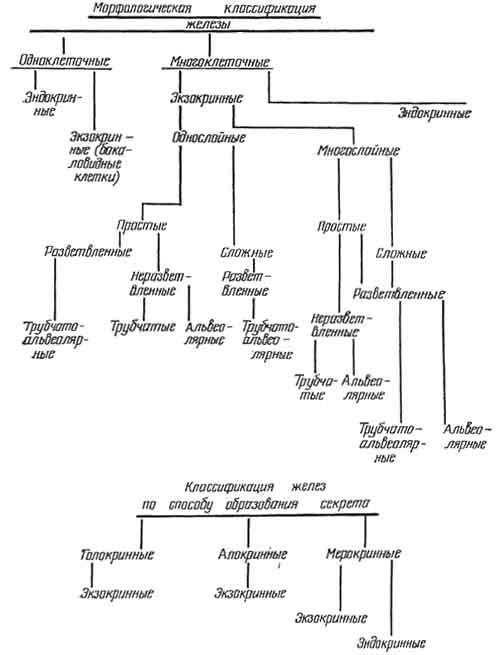
Рис. 92. Классификация желез.
Концевые отделы у простых желез разветвляются и не разветвляются, у сложных - разветвляются. В связи с этим у них и соответствующие названия: разветвленная железа и неразветвленная железа.
По форме концевых отделов экзокринные железы классифицируют на альвеолярные, трубчатые, трубчато-альвеолярные. У альвеолярной железы клетки концевых отделов формируют пузырьки или мешочки, у трубчатых - образуют вид трубочки. Форма концевой части трубчато-альвеолярной железы занимает промежуточное положение между мешочком и трубочкой (рис. 92, 93).
117

Рис. 93. Схематическое изображение простых и сложных экзокринных желез:
1 - простые трубчатые железы с неразветвленными концевыми отделами; 2 - простая альвеолярная железа с неразветвленным концевым отделом; 3 - простые трубчатые железы с разветвленными концевыми отделами; 4 - простые альвеолярные железы с разветвленными концевыми отделами; 5 - сложная альвеолярно-трубчатая железа с разветвленным концевым отделом; 6 - сложная альвеолярная железа с разветвленными концевыми отделами. Секреторные отделы изображены черными линиями, выводные протоки - светлыми.
Клетки концевого отдела именуются г л а н д у л о ц и т а м и . Процесс синтеза секрета начинается с момента поглощения гландулоцитами из крови и лимфы исходных компонентов секрета. При активном участии органелл, синтезирующих секрет белкового или углеводного характера, в гландулоцитах образуются секреторные гранулы. Они накапливаются в апикальной части клетки, а затем путем обратного пиноцитоза выделяются в полость концевого отдела. Завершающий этап секреторного цикла - восстановление клеточных структур, если в процессе секреции они разрушились.
Строение клеток концевой части экзокринных желез обусловлено составом выделяемого секрета и способом его образования.
По способу образования секрета железы делят на голокринные, апокринные, мерокринные (эккринные). При г о л о к р и н н о й секреции (HOLOS - целый) железистый метаморфоз гландулоцитов начинается с периферии концевого отдела и протекает в направлении выводного протока. Примером голокринной секреции является сальная железа. Стволовые клетки с базофильной цитоплазмой и округлым ядром расположены на периферии концевой части. Они интенсивно делятся митозом, поэтому мелкие по размеру. Перемещаясь к центру железы, секреторные клетки увеличиваются, так как в их цитоплазме постепенно накапливаются капельки кожного жира. Чем больше откладывается в цитоплазме жировых капель, тем интенсивнее протекает процесс деструкции органелл. Он завершается полным разрушением клетки. Плазмолемма разрывается, а содержимое гландулоцита поступает в просвет выводного протока.
При а п о к р и н н о й секреции (аро - от, сверху) разрушается апикальная часть секреторной клетки, являясь затем составной частью ее секрета. Данный тип секреции совершается в потовой или молочной железах.
При м е р о к р и н н о й секреции клетка не разрушается. Такой способ образования секрета типичен для многих желез организма: железы желудка, слюнные железы, поджелудочная железа, эндокринные железы (рис. 94).
118

Рис. 94. Различные тппы секреции:
А - мерокринный; F - апокринный; В - голокринный; 1 - малодифференцированные клетки; 2 - перерождающиеся клетки; 3 - разрушающиеся клетки.
Таким образом, железистый эпителий так же, как и покровный, развивается из всех трех зародышевых листков (эктодермы, мезодермы, энтодермы), расположен на соединительной ткани, лишен кровеносных сосудов, поэтому питание осуществляется диффузионным способом. Клеткам свойственна полярная дифференцировка: в апикальном полюсе локализуется секрет, в базальном полюсе - ядро и органеллы.
Регенерация. Покровные эпителии занимают пограничное положение. Они часто повреждаются, поэтому характеризуются высокой регенерационной способностью. Регенерация осуществляется главным образом митомическим и очень редко амитотическим способом. Клетки эпителиального пласта быстро изнашиваются, стареют и гибнут. Их восстановление называется ф и з и о л о г и ч е с к о й р е г е н е р а ц и е й .
Восстановление эпителиальных клеток, утраченных по причине травмы и другой патологии, называется р е п а р а т и в н о й р е г е н е р а ц и е й .
Воднослойных эпителиях регенерационной способностью обладают или все клетки эпителиального пласта, или, если эпптелиоциты высокодифференцированны, то за счет зонально лежащих своих стволовых клеток.
Вмногослойных эпителиях стволовые клетки находятся на базальной мембране, поэтому лежат в глубине эпителиального пласта.
Вжелезистом эпителии характер регенерации обусловлен способом образования секрета. При голокринной секреции стволовые клетки находятся снаружи железы на базальной мембране. Делясь и дифференцируясь, стволовые клетки преобразуются в железистые.
Вмерокринных и апокринных железах восстановление эпителиоцитов протекает главным образом путем внутриклеточной регенерации.
119
116 :: 117 :: 118 :: 119 :: Содержание

120 :: 121 :: Содержание
ТКАНИ ВНУТРЕННЕЙ СРЕДЫ (ОПОРНОТРОФИЧЕСКИЕ ТКАНИ)
Ткани внутренней среды возникают одновременно с эпителиальными на самых ранних этапах развития многоклеточных животных. У высших позвоночных они представлены группой тканей, общим морфологическим признаком которых является наличие в их составе не только клеток, но и хорошо развитого межклеточного вещества. В соответствии со специфичностью и различием клеточного состава и в большей степени особенностями структурной организации межклеточного вещества среди тканей внутренней среды выделяют: кровь и лимфу, разновидности соединительных тканей, хрящевую и костную ткани. Проявлением единства этих видов тканей при резком различии физико-химических свойств (кровь и лимфа - жидкие, костная ткань - твердая) является происхождение их из общего эмбрионального источника - мезенхимы.
Всем тканям внутренней среды свойственны трофическая и защитная, а тканям соединительным, хрящевым и костным - в той или иной степени механическая и опорная функции.
МЕЗЕНХИМА
Мезенхима - совокупность эмбриональных сетевидно связанных отростчатых клеток, заполняющих промежутки между более компактными эпителиоподобными зародышевыми листками и зачатками органов. В ячеях этой сети находится студенистое межклеточное вещество (рис. 95).
Рис. 95. Мезенхима.
120
При эмбриогенезе раньше всего мезенхима появляется в составе внезародышевых органов. Подтверждается это тем фактом, что первые кровяные островки возникают в

стенке желточного мешка. В теле зародыша мезенхима возникает главным образом из клеток определенных участков мезодермы - дерматомов, склеротомов и спланхнотомов. В области головы часть мезенхимы развивается из клеток, выселяющихся из эктодермальной ганглиозной пластинки, - нейромезенхима. Клетки мезенхимы быстро делятся митозом. В различных ее участках возникают многочисленные мезенхимные производные - кровяные островки с их эндотелием и клетками крови, клетки соединительных тканей и гладкой мышечной ткани, образуются уплотненные клеточные зачатки скелетных тканей и др.
121
120 :: 121 :: Содержание