

цитология гистология и эмбриология александровская козлов радостина
.pdfгранулярная шэдоплазматическая сеть; 11 - агранулярная эндоплазматическая сеть (ув. 46000); Г - 12 - базальная мембрана (ув. 46000). Стрелками показаны пиноцитозные пузырьки.
184
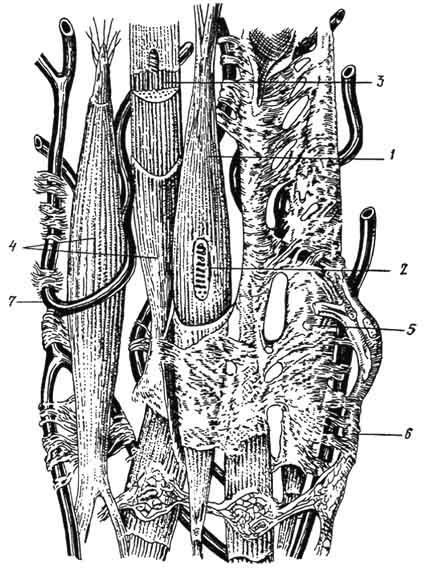
Рис. 136. Гладкомышечные клетки (тангенциальный срез):
A - 1 - ядро; 2 - цитоплазма; 3 - митохондрии; 4 - плазмолемма; 5 - толстые миофиламенты; 6 - коллаген; 7 - нервное волокно (ув. 14 000); Б - 8 - точки сближения плазмолемм двух соседних миоцитов (ув. 20 000); В - 9 - внутренняя мембрана митохондрии; 10 - наружная мембрана митохондрии; 11 - кристы; 12 - матрикс; 13 - миофиламенты (ув. 75 000). Электронная микрофотография.
185
Рис. 137. Схема строения гладкомышечного пласта:
1 - клетка мышечной ткани; 2 - ядро; 3 - миофиламенты: 4 - сарколемма; 5 - эндомизий; 6 - нерв; 7 - кровеносный капилляр.
незначительно выражены рибосомы, зернистая эндоплазматическая сеть (рис. 135). В цитоплазме миоцитов содержатся гранулы гликогена - энергетический резерв клетки.
Сократительным аппаратом миоцита являются миофиламенты. Они расположены в цитоплазме на периферии клетки и имеют продольную ориентацию. Новыми цитологическими методами выявлены три их разновидности: миозиновые (толстые), актиновые (тонкие) и промежуточные. Диаметр миозиновых миофиламентов равен 17 нм, актиновых - 7 нм, промежуточных - 10 нм. Миофиламенты построены из сократительных белков. В световой микроскоп миофиламенты неразличимы.
186
В гладкомышечных клетках отсутствует поперечная исчерченность. Это объясняется ориентацией миофиламентов.

Плазмолемма миоцита, впячиваясь в цитоплазму, формирует многочисленные округлые образования, сходные с пиноцитозными пузырьками (рис. 136), примыкающие к саркоплазматической сети. Предполагают, что они участвуют в проведении нервных импульсов к сократительному аппарату миоцита, обусловливая этим сокращение. В основе последнего, очевидно, лежит, как и в исчерченной мышечной ткани, процесс скольжения актиновых и миозиновых (миофиламентов) нитей по отношению друг к другу.
Снаружи миоциты покрыты базальной мембраной. Из коллагеновых и эластических волокон вокруг каждого миоцита формируется соединительнотканный футляр (эндомизий), который обеспечивает соединение миоцитов, не препятствуя процессу сокращения (рис. 137).
Если в мышечном пласте слабо развита соединительная ткань, то миоциты соединяются при помощи специальных межклеточных соединений типа десмосом (пятен слипания). Мышечный пласт построен так, что каждый его миоцит контактирует с несколькими соседними клетками. Такое расположение способствует более быстрому распространению нервного импульса, при этом в процесс сокращения вовлекается целая группа мышечных клеток.
Гладкомышечный пласт снабжен кровеносными, лимфатическими сосудами, нервными волокнами и окончаниями. В стенках многих внутренних полых органов гладкомышечные пласты формируют мышечные оболочки.
187
182 :: 183 :: 184 :: 185 :: 186 :: 187 :: Содержание

187 :: 188 :: 189 :: 190 :: 191 :: 192 :: 193 :: 194 :: 195 :: 196 :: 197 :: 198 :: 199 :: Содержание
ИСЧЕРЧЕННЫЕ (ПОПЕРЕЧНОПОЛОСАТЫЕ) МЫШЕЧНЫЕ ТКАНИ
Скелетная мышечная ткань - это сократительная ткань туловища, головы, конечностей, глотки, гортани, верхней половины пищевода, языка, жевательных мышц. Данную ткань относят к произвольной мускулатуре, так как ее сокращение контролируется волей животного.
Скелетная мышечная ткань развивается из миотомов сегментированного отдела мезодермы, а исчерченная мышечная ткань внутренних органов - из спланхнотома.
На ранней стадии развития миотомы состоят из плотно расположенных мышечных клеток - м и о б л а с т о в . Это первая стадия гистогенеза - м и о б л а с т и ч е с к а я . Цитоплазма миобластов имеет тонковолокнистое строение, свидетельствующее о развитии сократительных белков. Уже на этой стадии миобласты способны к сокращению. Они интенсивно делятся и перемещаются клеточными потоками в участки расположения будущих мышц (рис. 138). Вскоре в цитоплазме миобластов можно различить единичные сократительные нити - миофибриллы, построенные из сократительных белков. Ядра миобластов относительно крупные, овальные, с малым количеством гетерохроматина и
187
Рис. 138. Дифференцировка миобластов (т) в потоке клеток, выселяющихся из миотома.
хорошо выраженными ядрышками. Они делятся интенсивнее клеток, поэтому вскоре миобласты становятся многоядерными. Увеличиваясь в длину, они приобретают форму волокон - с и м п л а с т о в .
В центре симпласта в ряд располагаются многочисленные ядра, на периферии интенсивно дифференцируются миофибриллы. Миосимпласты, очевидно, могут
образовываться и путем слияния миобластов. Это вторая стадия гистогенеза. Она называется с т а д и е й м ы ш е ч н ы х т р у б о ч е к . Мышечные трубочки, расщепляясь вдоль, формируют мышечные волокна. В последних число миофибрилл резко увеличивается, многочисленные ядра перемещаются на периферию и располагаются под
плазмолеммой. Волокно |
приобретает исчерченность. Это третья стадия |
гистогенеза - |
||
с т а д и я |
м ы ш е ч н |
ы х |
в о л о к о н . К мышечным волокнам |
подрастает |
соединительная ткань с кровеносными сосудами, нервами, дифференцируются нервные окончания. Соединительная ткань участвует в образовании наружной оболочки мышечного волокна и связывает между собой мышечные волокна (рис. 139).
Сведения о гистогенезе помогут понять строение скелетной мышечной ткани и разобраться в тех сложных изменениях, которые протекают в ней при физической нагрузке, тренировке, в условиях физиологической регенерации и патологии.
Процесс регенерации, протекающий в скелетной мышечной ткани, сходен с гистогенезом; в нем выявлены те же
188
миобластическая стадия, стадия мышечных трубок и стадия мышечных волокон.
Как следует из гистогенеза, у дифференцированной скелетной мышечной ткани нет клеточного строения. Ее структурной и функциональной единицей является м ы ш е ч н о е в о л о к н о (рис. 140) в виде длинных цитоплазматических тяжей с закругленными концами, которые могут переходить в сухожилия. Длина волокон 10 - 100 мкм. Состоит мышечное волокно из саркоплазмы (цитоплазмы) и многочисленных ядер, расположенных на периферии. Само волокно покрыто сарколеммой (оболочкой). Структурные компоненты саркоплазмы - сократительный аппарат, органеллы, включения, гиалоплазма. Понять механизм сокращения скелетной мышечной ткани можно лишь после знакомства с тончайшей структурной организацией всех его компонентов.
Сократительным аппаратом скелетного мышечного волокна являются продольно ориентированные м и о ф и б р и л л ы . Построенные из сократительных белков, они занимают большую часть волокна, оттесняя ядра на периферию. Диаметр
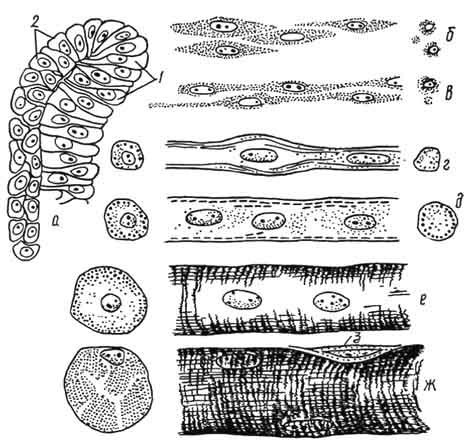
Рис. 139. Основные этапы эмбриогенеза скелетно-мышечной ткани:
а - клетки сомита (1 - миотом, 2 - дермотом); б - миобласты; в - миосимпласты; г - промиотуба; д - мышечная трубочка; е - незрелое мышечное волокно; ж - зрелое мышечное волокно; 3 - клетка соединительной ткани. Стадии б - ж показаны на продольном и поперечном разрезах.
189
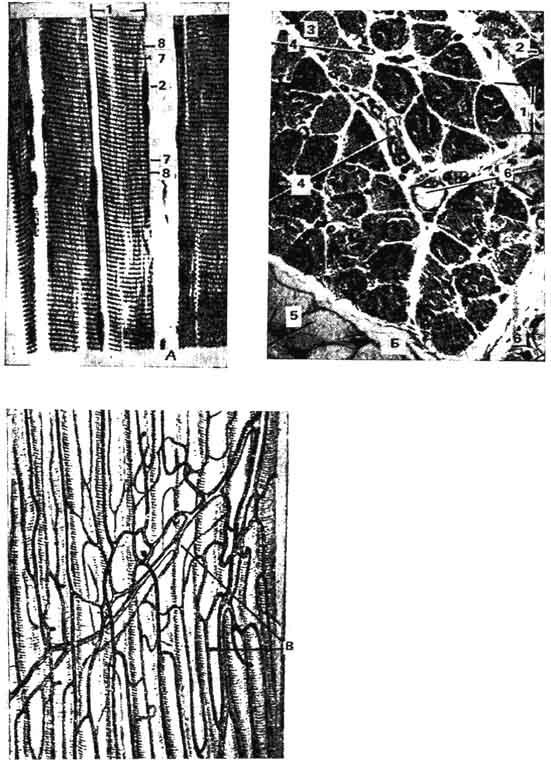
Рис. 140. Исчерченная скелетная мышечная ткань:
А - продольное сечение; Б - попереченое сечение; 1 - мышечное волокно; 2 - ядро мышечного волокна; 3 - миофибриллы; 4 - соединительная ткань перимизия; 5 - жировые клетки; 6 - кровеносный сосуд; 7 - анизотропный диск; 8 - изотропный диск; В - кровеносные сосуды мышечных волокон.
миофибрилл около 1 - 2 мкм. Миофибриллы состоят из чередующихся темных и светлых полос (дисков). Все светлые и все темные диски миофибрилл в одном мышечном волокне удерживаются на одном уровне, в связи с чем волокно приобретает поперечную исчерченность. Продольная ориентация миофибрилл

190
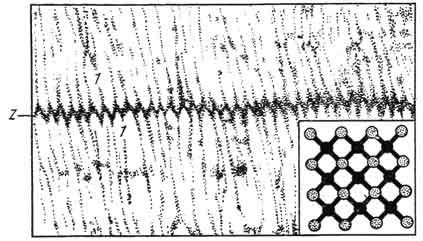
Рис. 141. Строение миофибрилл исчерченной скелетной мышечной ткани:
А - диск (анизотропный); I - диск (изотропный); Z-линия (телофрагма); М -линия (мезофрагма) (по Хаксли). Электронная микрофотография.
может создавать продольную исчерченность мышечного волокна.
В поляризованном свете темные полосы (диски) обнаруживают двойное лучепреломление - анизотропию, поэтому их называют анизотропными, или полосами А (дисками А). Светлые полосы изотропны, их называют изотропными, или полосами I (дисками I). В середине каждого I диска проходит темная зона - линия Z (телофрагма). В середине А диска проходит светлая зона - линия H с темной линией посередине - линией M (мезофрагма) (рис. 141), Диски и линии были открыты очень давно с помощью оптического микроскопа. Они хорошо видны на изолированных миофибриллах, которые можно получить, расщепив мышечное волокно.
Структурной единицей миофибриллы является саркомер. В миофибрилле они расположены, следуя друг за другом. Саркомер - это участок миофибржллы, состоящий из линия Z (для двух соседних саркомеров), половины диска I, диска А с линией Н, половины следующего диска I1 линии Z (для двух соседних саркомеров). Эти компоненты миофибрилл связывали с сокращениями, однако их участие в этом процессе оставалось неясным. Электронно-микроскопические, гистохимические, биохимические исследования многое внесли в расшифровку функциональной морфологии саркомера. Было установлено, что диск А состоит из более толстых (диаметром 10 нм, длиной 1,5 мкм) миофиламентов, диск I - из более тонких (диаметром 5 нм, длиной 1 мкм) миофиламентов. Материалом для построения толстых миофиламентов служит белок миозин, а тонких - актин, тропомиозин В, тропин.
Актиновые и миозиновые миофиламенты контактируют не конец в конец, а перемещаются по отношению друг к другу и в диске А образуют зону перекрытия. Участок А диска, состоящий только из миозиновых миофиламентов, называется H линией и по сравнению с зоной перекрытия более светлый. Линия M - это место соединения толстых миозиновых миофиламентов в анизотропном диске.
Линия Z состоит из Z-филаментов. В них выявлены белки тропомиозин-В, а-актин. Z- филаменты формируют решетку, к
191
PиC. 142. Линия Z:
1 - прикрепление к ней тонких миофиламентов. Вставка внизу поясняет прикрепление тонких миофиламентов к Z. Электронная микрофотография.
которой с обеих сторон прикреплены тонкие актиновые филаменты полосок I двух соседних саркомеров. Z линия проходит через всю толщину саркомера, а зона прикрепления тонких миофиламентов имеет зигзагообразный контур (рис. 142).
Таким образом, линии Z и M являются опорным аппаратом саркомера.
Встроении сократительного аппарата при сокращении мышечного волокна наблюдаются следующие изменения: уменьшается длина саркомеров, так как тонкие (актиновые) миофиламенты полоски I при скольжении между толстыми (миозиновыми) филаментами полоски А сдвигаются к линии M диска А. Это приводит к увеличению зоны перекрытия, формированию боковых мостиков между актиновыми и миозиновыми миофиламентами (рис. 143), сокращению линий H, сближению линий Z (рис. 144).
Вгиалоплазме мышечного волокна хорошо развиты митохондрии - органеллы клеточного дыхания. Они скапливаются между миофибриллами, вокруг многочисленных ядер, вблизи сарколеммы, то есть в тех зонах, которые характеризуются значительной потребляемостью АТФ. Этим объясняется высокая метаболическая активность скелетного мышечного волокна.
Интенсивное развитие в мышечном волокне имеет незернистая эндоплазматическая сеть (саркоплазматический ретикулум). Ее мембранные элементы расположены вдоль саркомеров и в виде терминальных цистерн окружают линии Z (рис. 145). Саркоплазматической сети присуща специфическая функция накопления ионов кальция, необходимых при сокращении и расслаблении мышечного волокна.
Остальные органеллы (зернистая эндоплазматическая сеть, комплекс Гольджи и др.) развиты слабее и локализуются около ядер.
192
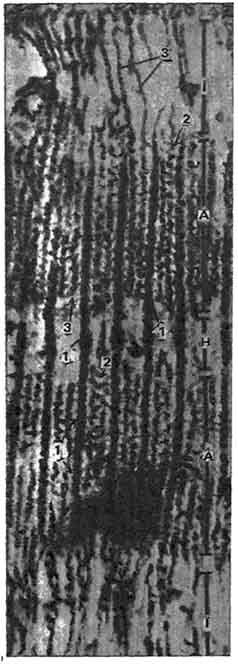
Рис. 143. Участок саркомера исчерченной мышечной ткани:
1 - толстые миофиламенты; 2 - поперечные мостики; 3 - тонкие миофиламенты. А - 1/2 диска A; I - 1/2 диска I; H - зона, состоящая только из толстых миофиламентов (по Хаксли).