

цитология гистология и эмбриология александровская козлов радостина
.pdfРис. 101. Электронная микрограмма пронормоцита (по Агееву):
1 - митохондрии; 2 - рибосомы; 3 - ядрышко.
140
базофильной цитоплазмой, в которой становится заметной узкая более светлая перинуклеарная зона - б а з о ф и л ь н ы е н о р м о ц и т ы .
Базофилия цитоплазмы обусловлена наличием в ней большого количества РНК, свободных рибосом и полисом, с которыми связан синтез белкового компонента гемоглобина. Последний накапливается сначала в перинуклеарной зоне. Железо, входящее в состав гемоглобина, базофильные нормоциты получают от макрофагов, фагоцитирующих гибнущие эритроциты. В результате деления базофильных нормоцитов появляются еще более мелкие клетки, цитоплазма которых из-за накопленного гемоглобина утрачивает выраженную ранее базофилию и окрашивается как основными, так и кислыми красителями - п о л и х р о м а т о ф и л ь н ы е н о р м о ц и т ы . Ядра их, как правило, без ядрышек, а вследствие радиального расположения грубых глыбок гетерохроматина имеют вид колеса со спицами. Проделав завершающее деление, полихроматофильные нормоциты превращаются в клетки, цитоплазма которых вследствие обилия в ней гемоглобина проявляет выраженную оксифилию - о к с и ф и л ь н ы е н о р м о ц и т ы . Ядро их постепенно уменьшается, пикнотизируется (уплотняется) и отсоединяется. Отделившиеся ядра нормоцитов фагоцитируются макрофагами костного мозга. Образуются молодые эритроциты - р е т и к у л о ц и т ы , поступающие в кровяное русло. В них еще некоторое время сохраняются РНК-содержащие структуры в виде сеточки. В процессе развития морфологически распознаваемые клетки эритроцитарного ряда осуществляют 5 - 6 митозов.
Установлено, что даже в нормальных условиях кроветворения часть эритробластов (до 10%) не завершает свой цикл развития до эритроцитов и с помощью макрофагов костного мозга разрушается.
Этот процесс, названный неэффективным эритропоэзом, является одним из физиологически обусловленных механизмов регуляции в системе эритропоэза. Наиболее сильным регулятором эритропоэза является количество кислорода, доставляемого к тканям и органам. Недостаточное снабжение кислородом стимулирует усиленную выработку эритропоэтина, посредством которого регулируется интенсивность пролиферации костномозговых предшественников (преимущественно на уровне БОЕ - Э и КОЕ - Э) эритропоэза. Эритропоэтиы - гормон гликопротеидной природы. Считают, что он синтезируется в почках.
Развитие гранулоцитов (гранулоцитопоэз). При развитии гранулоцитов из стволовых клеток красного костного мозга вначале также образуются морфологически нераспознаваемые полустволовые (КОЕ - ТЭММ; КОЕ - ГМ; КОЕ - ГЭ) и унипотентные предшественники (КОЕ - Б; КОЕ - Эо; КОЕ - Гн), которые через стадии распознаваемых клеточных форм (миелобласт, промиелоцит, миелоцит, метамиелоцит, палочкоядерный гранулоцит) превращаются в зрелые сегментоядерные гранулоциты трех разновидностей - нейтрофилы, эозинофилы и базофилы.
|
141 |
М и е л о б л а с т |
- первая морфологически идентифицируемая клетка |
гранулоцитарного ряда - имеет крупное центрально расположенное ядро, в котором на фоне диффузно-мелкозернистого хроматина видно несколько ядрышек. Цитоплазма слабобазофильная, в ней можно обнаружить небольшое количество азурофильных гранул. Электронно-микроскопически в ней выявляются митохондрии, полирибосомы, элементы гранулярной цитоплазматической сети. Разделившись, миелобласт превращается в п р о м и е л о ц и т - крупную клетку (20 - 25 мкм), в которой округлое ядро часто расположено эксцентрично и содержит 1 - 2 ядрышка. В элементах пластинчатого комплекса Гольджи промиелоцитов происходит основной процесс формирования азурофильных гранул. Они в значительном количестве содержатся в цитоплазме и дают положительную реакцию на пероксидазу. После деления из промиелоцита образуются клетки еще меньших размеров и с более плотным ядром, имеющим чаще всего овальную форму, - миелоциты. В цитоплазме миелоцитов наряду с первичной (азурофильной) зернистостью образуются и содержатся вторичные (специфические) гранулы, в соответствии с особенностями которых удается отчетливо различать нейтрофильные, эозинофильные и базофильные миелоциты.
Вразвитии гранулоцитов миелоциты являются завершающими клетками, способными
кмитотическому делению, после которого они созревают, последовательно превращаясь в метамиелоциты, палочкоядерные и сегментоядерные клетки. Эти этапы созревания характеризуются некоторыми общими морфологическими изменениями: уменьшением клетки в размерах, изменением формы ядра от овальной (миелоциты) через бобовидную (метамиелоциты) и палочковидную к расчлененной на дольки (сегментоядерные) и окончательным оформлением в цитоплазме соответствующей специфической зернистости. Сегментоядерные гарнулоциты поступают из костного мозга в кровяное русло, циркулируют в нем 8 - 12 ч, после чего проникают в ткани, где выполняют специфические функции и погибают. Показано, что для эозинофилов тканевая фаза их жизни продолжается около 10 сут, базофилы же погибают очень быстро.

Развитие моноцитов (моноцитопоэз). Клетки моноцитарпого ряда образуются в костном мозге из стволовых клеток через стадии полустволовых клеток (КОЕ - ГЭММ и КОЕ - ГМ), из которых возникают унипотентный предшественник (КОЕ - M) и затем монобласты. Немногочисленные монобласты трудно отличимы от других бластных форм в красном костном мозге. После того как в их цитоплазме сформируются компоненты комплекса Гольджи и образуются азурофильные гранулы, клетки превращаются в промоноциты и моноциты. Последние выходят в кровоток, а затем, проникая в ткани, дают начало незрелым, а позднее зрелым макрофагам.
Последовательное развитие клеток линии моноцитов - макрофагов из монобластов представлено на схеме.
142
Развитие лимфоцитов. Лимфоцитопоэз - один из наиболее сложных процессов дифференцировки стволовых кроветворных клеток. Важная особенность этого процесса состоит в том, что развивается сходная морфологически, но разнородная в функциональном отношении клеточная популяция. С участием различных органов поэтапно осуществляется формирование двух тесно связанных при функционировании линий клеток - T- и В- лимфоцитов. В красном костном мозге образуются родоначальные лимфоидные клетки, общие как для T-, так и для В-лимфоцитов. В центральных лимфоидных органах (тимусе, фабрициевой сумке) лимфоцитопоэз зависит от наличия жизнеспособных костномозговых предшественников. В периферических лимфоидных органах (лимфатические узлы, селезенка, лимфоидные образования слизистых оболочек) лимфоцитопоэз является антигензависимым процессом.
Развитие кровяных пластинок (тромбоцитопоэз). Образование кровяных пластинок происходит в красном костном мозге и связано с развитием в нем особых гигантских клеток - м е г а к а р и о ц и т о в . Мегакариоцитопоэз состоит из следующих стадий: стволовая клетка (CK) - полустволовые клетки (КОЕ - ГЭММ и КОЕ - МГЦЭ) - унипотентные предшественники (КОЕ - МГЦ) - мегакариобласт - промегакариоцит - мегакариоцит. По мере созревания, в результате своеобразной многократной эндометотпческой репродукции, формируются крупные клетки (40 - 50 мкм), содержащие в многолопастном ядре до 32 - 64 хромосомных наборов. В развивающихся мегакариоцитах, в цитоплазме образуются система микрофиламентов и микротрубочек, а также специфические гранулы. На заключительных этапах с участием формирующейся системы из гладких мембран происходит фрагментация цитоплазмы мегакариоцитов на обособленные участки - кровяные пластинки, которые через стенки синусоидов красного костного мозга попадают в кровоток. После отсоединения пластинок вокруг оставшегося ядра мегакариоцитов возникает новая цитоплазма. Полагают, что в каждом мегакариоците красного костного мозга совершается циклический процесс развития нескольких поколений кровяных пластинок.
143
137 :: 138 :: 139 :: 140 :: 141 :: 142 :: 143 :: Содержание

143 :: 144 :: 145 :: Содержание
Эмбриональное кроветворение
В течение эмбрионального периода развития организма происходит последовательная смена локализации кроветворения. На самых ранних этапах клетки крови образуются за пределами зародыша, в мезенхиме стенки желточного мешка, где формируются скопления - к р о в я н ы е о с т р о в к и . Центральные клетки островков округляются, обособляются и преобразуются в стволовые кроветворные клетки. Периферические клетки островков
143
растягиваются в плоские, связанные между собой клетки и образуют эндотелиальную выстилку первичных кровеносных сосудов. В результате соединения разрозненных вначале островков образуется сосудистая сеть желточного мешка.
Часть стволовых клеток превращается в крупные базофильные бластные клетки - п е р в и ч н ы е к р о в я н ы е к л е т к и . Интенсивно размножаясь внутри сосуда, большая часть первичных кровяных клеток постепенно утрачивает базофилию, все сильнее окрашиваясь кислыми красителями. Это происходит в связи с синтезом и накоплением в цитоплазме гемоглобина. Одновременно в ядре увеличивается количество конденсированного хроматина. Такие клетки называют п е р в и ч н ы м и э р и т р о б л а с т а м и . В некоторых первичных эритробластах распадается и исчезает ядро. Образующаяся генерация ядерных и безъядерных первичных эритроцитов
разнообразна по размерам, однако чаще |
всего встречаются |
крупные клетки - |
м е г а л о б л а с т ы и м е г а л о ц и т ы . |
Мегалобластический |
тип кроветворения |
характерен для эмбрионального периода. При некоторых формах анемий (В12-дефицитные, фолиеводефицитные и др.) в крови взрослых животных появляются клетки, сходные с первичными мегалоцитами.
Часть первичных кровяных клеток в эмбриональных сосудах проходит более длительный путь развития и образует популяцию вторичных эритроцитов, имеющих меньшие размеры и по внешнему виду мало отличающихся от эритроцитов взрослых животных.
Одновременно с внутрисосудистым эритропоэзом из первичных кровяных клеток вне сосудов развивается небольшое количество гранулоцитов - нейтрофилов и эозинофилов, т. е. происходит миелопоэз.
Возникшие в желточном мешке стволовые клетки с кровью переносятся в различные формирующиеся органы организма, где и происходит их дальнейшее развитие в разнообразные клетки крови. После закладки печени она становится универсальным органом кроветворения; в ней экстраваскулярно развиваются вторичные эритроциты, зернистые лейкоциты и мегакариоциты. К концу внутриутробного периода кроветворение в печени прекращается.
Из стволовых клеток в развивающемся тимусе (на 7 - 8-й неделе у крупного рогатого скота) дифференцируются лимфоциты тимуса и мигрирующие из него Т-лимфоциты. Последние заселяют Т-зоны развивающихся периферических кроветворных органов - селезенки, лимфатических узлов. В начале своего развития селезенка также является органом, в котором образуются все виды форменных элементов крови. У большинства животных к концу эмбрионального периода в связи с массовым вселением T- и B-

лимфоцитов из центральных органов в селезенке и лимфатических узлах начинает преобладать лимфоцитопоэз.
На поздних стадиях эмбрионального развития у животных основные кроветворные функции (роль центрального органа) начинает выполнять красный костный мозг; в нем образуются эритроциты, гранулоциты, кровяные пластинки, часть лимфоцитов
144
(В-лимфоциты). В красном костном мозге сохраняется основная масса самоподдерживающей популяции стволовых клеток, и он становится органом, где происходит универсальный гемопоэз в постэмбриональный период. Таким образом, кроветворение, присущее всей мезенхиме, становится функцией специализированных органов (печени, селезенки, тимуса, красного костного мозга, лимфатических узлов), где создаются соответствующие условия микроокружения и кровотока.
Во время эмбрионального эритроцитопоэза идет характерны II процесс смены генераций эритроцитов, отличающихся морфологией и типом образующегося гемоглобина. Популяция первичных эритроцитов образует эмбриональный тип гемоглобина (HB - Г). На последующих стадиях эритроциты в печени и селезенке содержат плодный (фетальный) тип гемоглобина (HB - Г). Наконец, на смену им появляется дефинитивный тип эритроцитов, которые образуются в красном костном мозге, они содержат третий тип гемоглобина (HB - А и HB - А2). Разные типы гемоглобинов отличаются составом аминокислот в белковой части.
145
143 :: 144 :: 145 :: Содержание

145 :: 146 :: 147 :: 148 :: 149 :: 150 :: 151 :: 152 :: 153 :: 154 :: 155 :: 156 :: 157 :: 158 :: 159 :: 160 :: 161 :: 162 ::
Содержание
СОЕДИНИТЕЛЬНЫЕ ТКАНИ
Широко распространенные в организме животных ткани с сильна развитой в межклеточном веществе системой волокон, благодаря которым эти ткани выполняют разносторонние механические и формообразующие функции - формируют комплекс перегородок, трабекул или прослоек внутри органов, входят в состав многочисленных оболочек, образуют капсулы, связки, фасции и сухожилия.
В зависимости от количественного соотношения между компонентами межклеточного вещества - волокнами и основным веществом и в соответствии с типом волокон различают тря вида соединительных тканей: рыхлую соединительную ткань, для которой характерно количественное преобладание основного вещества над комплексом разнообразно ориентированных и рыхло расположенных коллагеновых и эластических волокон; плотную соединительную ткань, в ней резко выражено преобладание волокон над основным веществом, и ретикулярную ткань, содержащую в своем составе специфические ретикулярные волокна.
Основными клетками, создающими вещества, необходимые для построения волокон в рыхлой и плотной соединительной ткани, являются фибробласты, в ретикулярной ткани - ретикулярные клетки. Рыхлая соединительная ткань отличается особенно большим разнообразием клеточного состава.
РЫХЛАЯ СОЕДИНИТЕЛЬНАЯ ТКАНЬ
В организме рыхлая соединительная ткань является наиболее распространенной. Об этом свидетельствует тот факт, что она в большем или меньшем количестве сопровождает все кровеносные
145
и лимфатические сосуды, формирует многочисленные прослойки внутри органов, входит в состав кожи и слизистых оболочек внутренних полостных органов.
Независимо от локализации рыхлая соединительная ткань состоит из разнообразных клеток и межклеточного вещества, содержащего основное (аморфное) вещество и систему коллагеновых и эластических волокон. В соответствии с местными условиями развития и функционирования количественное соотношение между этими тремя структурными элементами в различных участках неодинаковое, что обусловливает органные особенности рыхлой соединительной ткани.
Среди разнообразных высокоспециализированных клеток в ^составе данной ткани различают более оседлые клетки (фибробласты - фиброциты, липоциты), развитие которых в процессе клеточного обновления происходит из предшественников, расположенных в пределах самой рыхлой соединительной ткани. Непосредственными предшественниками других более подвижных клеток (гистиоцпты - макрофаги, тканевые базофилы, плазмоциты) являются клетки крови, активная фаза функционирования которых осуществляется в составе рыхлой соединительной ткани. В совокупности все клетки рыхлой соединительной ткани представляют единый диффузно рассредоточенный аппарат,
находящийся в неразрывной функциональной связи с клетками сосудистой крови и лимфоидной системы организма.
Повсеместная распространенность рыхлой соединительной ткани, разнообразие и большое количество клеточных элементов, способных к размножению и миграции, обеспечивает главные функции этой соединительной ткани: трофическую (обменные процессы и регуляция питания клеток), защитную (участие клеток в иммунных реакциях - фагоцитоз, выработка иммуноглобулинов и других веществ) и пластическую (участие в восстановительных процессах при тканевом повреждении).
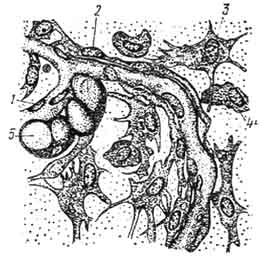
К л е т к и . Адвентициальные клетки - вытянутые клетки звездчатой формы с овальным ядром, богатым гетерохроматином. Цитоплазма базофильна, содержит немногочисленные органеллы. Располагаются вдоль наружной поверхности стенки капилляров и являются относительно малодифференцированными клеточными элементами, способными к митотическому делению и превращению в фибробласты, миофибробласты и липоциты (рис. 102).
Фибробласты (fibra - волокно, blastos - росток, зачаток) - - постоянные и наиболее многочисленные клетки всех видов соединительной ткани. Это основные клетки, принимающие непосредственное участие в формировании межклеточных структур. Они синтезируют и выделяют высокомолекулярные вещества, необходимые как для построения волокон, так и для образования аморфного компонента ткани. В ходе зародышевого развития фибробласты возникают непосредственно из мезенхимных клеток. В постэмбриональном периоде и при регенерации основными предшественниками фибробластов являются адвентициальные
146
Рис. 102. Рыхлая соединительная ткань подкожной клетчатки кролика (по Ясвоину):
1 - эндотелий капилляра; 2 - адвентициальная камбиальная клетка; 3 - фибробласты; 4 - гистиоцит; 5 - жировая клетка.
клетки. Кроме того, молодые формы этих клеток сохраняют способность размножаться путем митотического деления.
По степени зрелости и, следовательно, по структурной характеристике и функциональной активности различают три разновидности фибробластов. М а л о д и ф ф е р е н ц и р о в а н н ы е фибробласты имеют вытянутую, веретеновидную

форму с немногочисленными короткими отростками. Овальное ядро имеет хорошо выраженное ядрышко. Цитоплазма при световой микроскопии препаратов, окрашенных основными красителями, базофильная. При электронной микроскопии в цитоплазме обнаруживают много свободных полисом и лишь короткие узкие канальца гранулярной сети. Элементы комплекса Гольджи расположены в околоядерной зоне. Митохондрии немногочисленны и имеют плотный матрикс. Считают, что такие незрелые клетки обладают низким уровнем синтеза специфических белков. Их функция сводится к синтезу и секреции гликозаминогликанов.
З р е л ы е ф и б р о б л а с т ы при рассматривании сверху - крупные (диаметр до 50 мкм) отростчатые клетки, содержат светлые овальные ядра с 1 - 2 крупными ядрышками и значительный объем слабобазофильной цитоплазмы. Периферическая зона клетки окрашивается особенно слабо, вследствие чего контуры ее почти незаметны. В поперечном разрезе уплощенное тело клетки, веретеновидной формы, так как его центральная часть, содержащая ядро, значительно утолщена. Электронно-микроскопически для цитоплазмы зрелого фибробласта характерна развитая гранулярная эндоплазматическая сеть, состоящая из удлиненных и расширенных цистерн, к мембранам которых прикреплены полисомы больших размеров. Хорошо выражены и элементы (цистерны, микро- и макропузырьки) комплекса Гольджи, распределенные по всей цитоплазме. Обнаруживаются различные по форме и величине митохондрии (рис. 103).
В функциональном отношении зрелые фибробласты являются клетками со сложной синтетической и секреторной деятельностью. Они одновременно синтезируют и выводят несколько типов специфических белков (проколлаген, проэластин, ферментные белки) и разнообразные гликозаминогликаны. Наиболее отчетливо выражена способность к синтезу белка коллагеновых волокон. На
147
Рис. 103. Электронная микрограмма участка фибробласта (по Радостиной):
1 - ядро; 2 - гранулярная эндоплазматическая сеть; 3 - митохондрии.
полисомах гранулярной сети синтезируются α-цепи белка, а в полости компонентов ретикулума осуществляется их связывание в тройную спираль молекулы проколлагена. Последние с помощью микропузырьков переносятся в цистерны комплекса Гольджи и затем в составе секреторных гранул выделяются из клетки. На поверхности фибробласта от молекул проколлагена отделяются концевые неспирализованные пептидные участки, они превращаются в молекулы тропоколлагена, которые, полимеризуясь, формируют коллагеновые микрофибриллы и фибриллы (рис. 104). Гликозаминогликаны образуются в комплексе Гольджи. Накапливаясь между клетками, они создают условия для концентрации и полимеризации молекул тропоколлагена, а также входят в состав фибрилл в качестве цементирующего компонента.
Фибробласты обладают подвижностью. В периферической зоне цитоплазмы располагаются актинсодержащие микрофиламенты, при сокращении которых обеспечиваются образование выступов и движение клетки. Двигательная активность фибробластов усиливается в регенераторную фазу воспалительной реакции при образовании соединительнотканной капсулы.
В грануляционной ткани появляются фибробласты с большим количеством сократимых нитей - м и о ф и б р о б л а с т ы , способствующие закрытию раны.
Волокнообразование в соединительной ткани приводит к тому, что часть фибробластов оказывается заключенной между близко расположенными волокнами. Такие клетки называют ф и б р о ц и т а м и . Они утрачивают способность к делению, принимают сильно вытянутую форму, у них уменьшается объем цитоплазмы и значительно снижается синтетическая активность.
Гистиоциты (макрофаги) в составе широко распространенной соединительной ткани являются наиболее многочисленной группой свободных, способных к миграции клеток, относящихся к системе мононуклеарных фагоцитов (СМФ). В соединительнотканных прослойках разных органов их количество неодинаковое и, как правило, значительно увеличивается при воспалении.
148

Pиc. 104. Схема образования коллагеновой фибриллы:
А - аминокислоты (пролин, лизин и др.), поглощенные фибробластом, включаются в белок, синтезируемый на рибосомах эндоплазматической сети. Белок поступает в комплекс Гольджи, а затем выводится из клетки в форме молекул тропоколлагена, из которых вне клетки образуются коллагеновые фибриллы; 1 - фибробласт; 2 - гранулярная эндоплазматическая сеть; 3 - комплекс Гольджи; 4 - митохондрии; 5 - молекулы тропоколлагена; 6 - коллагеновая фибрилла (по Вельшу и Шторху).
При световой микроскопии окрашенных пленочных препаратов гистиоциты имеют разнообразную форму и разную величину (10 - 50 мкм), располагаются одиночно или группами. В сравнении с фибробластами выделяются более очерченными, но неровными границами и интенсивно окрашенной цитоплазмой, в которой имеются вакуоли и включения. Ядро небольшое, овальное, слегка вогнутое, содержит много глыбок гетерохроматина, в связи с чем оно более темное (рис. 105).
Электронно-микроскопически на плазмолемме отмечают микроворсинки, псевдоподии, инвагинации. В цитоплазме присутствует значительное количество лизосом, фагосом, гранул и липидных включений. Гранулярная сеть почти не развита. Митохондрии и комплекс Гольджи более развиты в активированных макрофагах. Цитохимическими методами в цитоплазме гистиоцитов выявляют разнообразные ферменты (кислые гидролазы, изоферменты кислой фосфатазы, эстеразы и др.), с помощью которых происходит переваривание поглощенных веществ.
П о н я т и е о с и с т е м е м о н о н у к л е а р н ы х ф а г о ц и т о в (макрофагической системе). К этой системе относят
149