

цитология гистология и эмбриология александровская козлов радостина
.pdfРис. 105. Рыхлая соединительная ткань в пленочном: препарате:
1 - фибробласты; 2 - гистиоциты; 3 - тканевый базофил; 4 - коллагеновые волокна; 5 - эластические волокна.
находящиеся во многих тканях и органах клетки, обладающие интенсивным эндоцитозом (фагоцитозом и пиноцитозом) макромолекулярных веществ экзогенной и эндогенной природы, частиц, микроорганизмов, вирусов, клеток, продуктов клеточного распада и т. п. Все макрофаги независимо от локализации происходят из стволовой кроветворной клетки красного костного мозга, а их непосредственными предшественниками являются моноциты периферической крови. Моноциты, вышедшие из сосуда и попавшие в соответствующее микроокружение, адаптируются к новой среде и превращаются в органо- и тканевоспецифические макрофаги (рис. 106).
Рис. 106. Разновидности клеток, принадлежащих к системе мононуклеарных фагоцитов - СМФ (по Ван-Фюрту, 1980).
150
Таким образом, моноциты циркулирующей крови представляют подвижную популяцию относительно незрелых клеток будущих зрелых макрофагов на их пути от костного мозга в органы и ткани. В условиях культивирования макрофаги способны прочна прикрепляться к поверхности стекла и приобретать уплощенную форму.
В зависимости от локализации (печень, легкие, брюшная полость и т. п.) макрофаги приобретают некоторые специфические черты строения и свойства, позволяющие отличать их друг от друга, однако всем им свойственны некоторые общие структурные, ультраструктурные и цитохимические признаки. Благодаря наличию сократимых микрофиламентов, обеспечивающих подвижность плазмолеммы, клетки этой системы способны к образованию различных приспособлений (ворсинок, псевдоподий, выступов), облегчающих захватывание частиц. Один из основных ультраструктурных признаков макрофагов - наличие в их цитоплазме многочисленных лизосом и фагосом. С участием лизосомальных ферментов (фосфатазы, эстеразы и др.) фагоцитированный материал расщепляется и перерабатывается.
Макрофаги - многофункциональные клетки. Основоположником учения о цитофизиологии клеток макрофагической системы .является И. И. Мечников. До настоящего времени актуальны многие положения, сформулированные им о механизмах фагоцитоза и биологическом значении этого явления. Макрофагическая система благодаря способности ее клеток поглощать и переваривать различные продукты экзо- и эндогенного происхождения представляет одну из важнейших защитных систем, участвующих в поддержании стабильности внутренней среды организма.
Макрофагам отведена важная роль в осуществлении защитной воспалительной реакции. Обладая способностью к направленному движению, определяемому хемотаксическими факторами (вещества, выделяемые бактериями и вирусами, иммунные комплексы антиген - антитело, продукты тканевого распада, медиаторы лимфоцитов и др.), макрофаги мигрируют в очаг воспаления и становятся доминирующими клетками хронического воспаления. При этом они не только очищают очаг от инородных частиц и разрушенных клеток, но и стимулируют в последующем функциональную активность фибробластов. При наличии в очаге токсичных и устойчивых раздражителей (некоторые микроорганизмы, химические вещества, малорастворимые материалы) с участием макрофагов формируется гранулема, в которой путем слияния клеток могут образовываться гигантские многоядерные клетки.
Макрофаги имеют существенное значение во многих иммунологических реакциях: в распознавании антигена, его переработке и предъявлении лимфоцитам, в межклеточном взаимодействии с T- и В-лимфоцитами и в выполнении эффекторных функций.
На поверхности плазмолеммы макрофага расположено два типа специфических рецепторов: рецепторы для Fc-части иммуноглобулинов д рецепторы для комплемента, особенно для его
151
компонента С3. Поэтому в фазе распознавания и поглощения большое значение имеет опсонизация антигенов, то есть предварительное присоединение к ним иммуноглобулинов или комплекса иммуноглобулина с комплементом. Последующее прикрепление таких сенсибилизированных антигенов (иммунных комплексов) к соответствующим рецепторам
макрофага вызывает движение псевдоподий и поглощение объекта фагоцитоза. Существуют также неспецифические рецепторы, благодаря которым клетка может фагоцитировать денатурированные белки или индифферентные частицы (полистирол, пыль и т. п.). С помощью пиноцитоза макрофаги способны распознавать и поглощать растворимые антигены (глобулярные белки и др.).
Большая часть поглощенных антигенных веществ во многих фагоцитах разрушается полностью. Такая функция ликвидации избытка антигенов, проникших во внутреннюю среду организма, характерна для макрофагов печени, синусов селезенки, мозгового вещества лимфатических узлов. Особые разновидности специализированных макрофагов - отростчатые "дендритные" клетки В-зон и "интердигитирующие" клетки Т-зон лимфатических узлов и селезенки. На поверхности их многочисленных отростков концентрируются и сохраняются исходные или частично переработанные иммуногенные антигены. В этих зонах макрофаги вступают в кооперативное взаимодействие с В- и Т- лимфоцитами про развитии как гуморального, так и клеточного иммунитета.
Эффекторная функция макрофагов выражается не только в обезвреживании антигенов, но и в способности вырабатывать и выделять растворимые факторы, стимулирующие пролиферацию и дифференциацию T- и В-лимфоцитов и регулирующие их активность. В некоторых случаях макрофаги могут быть эффекторными клетками в цитотоксических реакциях. Считают, что это важный механизм разрушения клеток с внутриклеточно паразитирующими микроорганизмами и опухолевых клеток.
Клетки системы мононуклеарных фагоцитов - активные участники миелоидного и лимфоидного кроветворения. Макрофаги в красном костном мозге являются своеобразными центрами, вокруг которых группируются предшественники развивающихся эритроцитов. Эти макрофаги участвуют в передаче накапливаемого железа в клетки эритроидного ряда, поглощают ядра нормоцитов и фагоцитируют поврежденные и старые эритроциты. Другие макрофаги костного мозга фагоцитируют части мегакариоцитов после отделения от них кровяных пластинок. С помощью макрофагов селезенки происходит интенсивный эритрофагоцитоз и поглощение стареющих кровяных пластинок, а макрофагов всех лимфоядных органов - фагоцитоз плазмоцитов и лимфоцитов.
Тканевые базофилы (лаброциты, тучные клетки) обнаружены у большинства позвоночных и у всех млекопитающих, однако количество их у животных разных видов и в соединительной ткани различных органов неодинаковое. У некоторых животных отмечают обратно пропорциональную зависимость между количеством
152
тканевых базофилов и базофилов крови, что свидетельствует о сходном биологическом значении этих типов клеток в системе тканей внутренней среды (например, у морских свинок тканевых базофилов много, но мало базофилов крови). Значительное количество тканевых базофилов содержится в подэпителиальной соединительной ткани кожи, пищеварительного тракта, дыхательных путей, матки. Их обнаруживают в соединительнотканных прослойках по ходу мелких кровеносных сосудов в печени, почках, эндокринных органах, молочной железе и других органах.
По форме тканевые базофилы чаще овальные или шаровидные, размером от 10 до 25 мкм. Ядро расположено центрально, в нем много глыбок конденсированного хроматина. Наиболее характерная структурная особенность тканевых базофилов - наличие многочисленных крупных (0,3 - 1 мкм) специфических гранул, равномерно заполняющих
большую часть объема цитоплазмы и окрашивающихся метахроматически. Электронномикроскопически в цитоплазме обнаруживают немногочисленные митохондрии, полисомы и рибосомы. Эндоплазматическая сеть и комплекс Гольджи развиты слабо. На плазмолемме пальцевидные выпячивания. Специфические гранулы окружены мембраной и имеют неодинаковую электронную плотность; в некоторых гранулах содержатся еще более электроноплотные зерна или пластинки.
Характерное метахроматическое окрашивание гранул обусловлено наличием в них сульфатированного гликозаминогликана - гепарина. Кроме того, в гранулах тканевых базофилов содержатся важнейшие биологические амины - гистамин, серотонин, дофамин, имеющие многообразное фармакологическое действие. Цитохимическими методами в цитоплазме выявлены различные ферменты - кислая и щелочная фосфатазы, липаза. Гистамин образуется из аминокислоты гистидина с помощью гистидиндекарбоксилазы, которая является маркерным ферментом для тучных клеток.
Располагаясь вблизи мелких кровеносных сосудов, тканевые базофилы одни из первых клеток реагируют на проникновение антигенов из крови. На их плазмолемме, так же как и у базофилов крови, находится значительное количество иммуноглобулинов класса E (IgE). Связывание антигенов и образование комплекса антиген - антитело сопровождается дегрануляцией и выделением из тканевых базофилов сосудистоактивных веществ, обусловливающих появление местных и общих реакций. Гистамин повышает проницаемость стенки капилляров и основного вещества соединительной ткани, стимулирует миграцию эозинофилов, активирует макрофаги и т. д. Гепарин препятствует свертыванию крови. Установлено участие тканевых базофилов при развитии аллергических и анафилактических реакций.
Дегрануляция тканевых базофилов может быть вызвана и различными физическими факторами - травмой, резкими температурными воздействиями и др.
Плазмоциты (плазматические клетки) в функциональном
153
отношении - эффекторные клетки иммунологических реакций гуморального типа, то есть реакции, сопровождающихся увеличением в крови циркулирующих антител, с помощью которых осуществляется обезвреживание антигенов, вызвавших их образование. Это высокоспециализированные клетки организма, синтезирующие и выделяющие основную массу разнообразных антител (иммуноглобулинов) .
По происхождению плазмоциты представляют конечные стадии развития стимулированных антигеном В-лимфоцитов. которые в местах своего расположения с участием клеток Т-хелперов в макрофагов подвергаются активации, интенсивно размножаются и превращаются в зрелые плазматические клетки. В наибольшем количестве плазмоциты находятся в селезенке, лимфатических узлах, в составе соединительной ткани слизистых оболочек пищеварительного канала и дыхательных путей, в интерстициальной соединительной ткани различных желез организма.
Развитие клеток плазматического ряда из стимулированных В-лимфоцитов происходит через стадии плазмобласта (иммунобласта), проплазмоцита и плазмоцита (рис. 107).
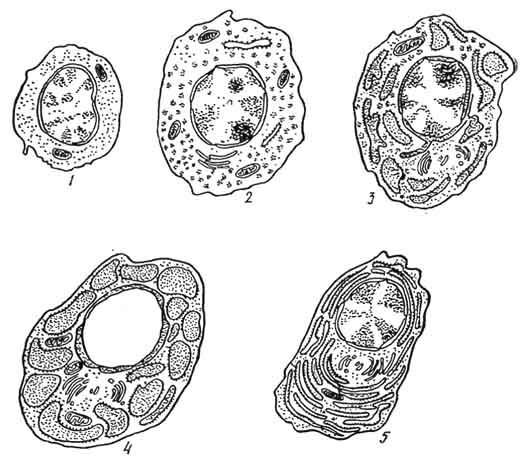
Рис. 107. Схема развития плазмоцитов (по Вайсу):
1 - предшественник плазмоцита (полустволовая клетка); 2 - плазмобласт; 3 - юный плазмоцит; 4 - плазмоцит с расширенными цистернами эндоплазматической сети; 5 - зрелый плазмоцит.
154
П л а з м о б л а с т - крупная клетка (до 30 мкм в диаметре), имеющая светлое центрально расположенное ядро. В последнем обнаруживают небольшие гранулы расположенного по периферии хроматина и 1 - 2 ярко выраженных ядрышка. Электронномикроскопически в цитоплазме выявляют очень редкие и мелкие цистерны гранулярной эндоплазматической сети, а также большое количество свободных полисом и рибосом. У немногочисленных митохондрий светлые матрикс и редкие кристы. Среди плазмобластов часто встречаются митозы. П р о п л а з м о ц и т характеризуется несколько меньшими размерами, резко выраженной базофилией цитоплазмы и неровной из-за многочисленных выпячиваний плазмолеммы поверхностью клетки. В цитоплазме находится большое количество расширенных цистерн и мешочков гранулярной эндоплазматической сети. Между элементами гранулярной сети расположены небольшие по размеру митохондрии. Считают, что эти клетки могут вырабатывать и выделять иммуноглобулины. З р е л ы е п л а з м о ц и т ы - сравнительно небольшие (8 - 10 мкм) клетки овальной формы с выраженными границами. В сильнобазофильной (пиронинофильной) цитоплазме обнаруживают светлую околоядерную зону. Ядро округлое, расположено эксцентрично и содержит крупные глыбки гетерохроматина, распределенные в виде спиц колеса. Особенно характерно в строении этих клеток при электронно-микроскопическом исследовании наличие в цитоплазме многочисленных, расположенных близко друг к другу длинных цистерн, имеющих очень узкую полость и сближенные мембраны, на внешней поверхности которых находятся многочисленные полисомы. В околоядерной зоне с более светлой
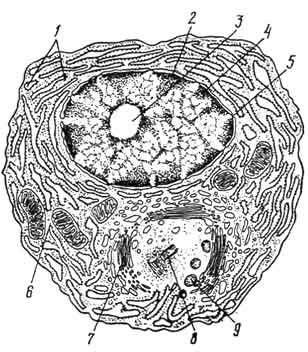
цитоплазмой этих цистерн нет, в ней расположены центриоли и хорошо развитый комплекс Гольджи (рис. 108).
Таким образом, в конечной стадии развития плазмоциты содержат мощный белоксинтезирующий аппарат, с помощью которого осуществляется синтез молекул иммуноглобулинов (антител). Установлено, что легкие цепи иммуноглобулинов синтезируются
Рис. 108. Схема ультрамикроскопического строения плазматической клетки (по Бесси):
1 - гранулярная эндоплазматическая сеть; 2 - хроматин; 3 - ядрышко; 4 - ядерная оболочка; 5 - пора ядерной оболочки; 6 - свободные рибосомы; 7 - комплекс Гольджи; 8 - центриоли; 9 - секреторные пузырьки.
155
на полирибосомах гранулярной сети отдельно от тяжелых цепей. Последние отделяются от полирибосом после образования их комплекса с легкими цепями. Так как весь белоксинтезирующий механизм запрограммирован на синтез антител только одной разновидности, каждая плазматическая клетка определенного клона способна за один час синтезировать несколько тысяч молекул иммуноглобулинов. Синтезированные молекулы поступают в просвет цистерн, а затем в комплекс Гольджи, откуда после присоединения углеводного компонента выводятся на поверхность клетки и выделяются. Выделение антител происходит и при разрушении клетки.
Превращение В-лимфоцита в плазмоцит длится около суток; продолжительность активной антителопродуцирующей деятельности зрелых плазмоцитов составляет несколько дней. Зрелые плазматические клетки не способны к делению, они стареют, погибают и поглощаются макрофагами.
Жировые клетки (липоциты) и жировая ткань (textus adiposus). Жировые клетки специализированы на синтезе и накапливании в цитоплазме запасных липидов, главным образом триглицеридов и утилизации их в соответствии с энергетическими и другими потребностями организма. Липоциты широко распространены в рыхлой соединительной ткани и чаще располагаются не одиночно, а небольшими группами по ходу мелких
кровеносных сосудов. Во многих частях организма животных образуются значительные скопления жировых клеток, называемые жировой тканью. В эмбриогенезе жировые клетки возникают из клеток мезенхимы. Предшественниками для образования новых жировых клеток в постэмбриональный период являются адвентициальные клетки, сопровождающие кровеносные капилляры.
В связи с особенностями естественной окраски клеток, спецификой их строения и функции, а также расположением различают у млекопитающих две разновидности жировых клеток и соответственно два типа жировой ткани: белую и бурую.
Б е л а я ж и р о в а я т к а н ь в организме животных разных видов и пород распределена неодинаково. В значительном количестве она содержится в жировых депо: подкожная жировая ткань, особенно развитая у свиней, жировая ткань вокруг почек, в брыжейке, у некоторых пород овец у корня хвоста (курдюк). У животных мясных и мясомолочных пород группы жировых клеток располагаются в перимизии и эндомизии внутри скелетных мышц. Мясо, полученное от таких животных, обладает наилучшими качествами ("мраморное" мясо).
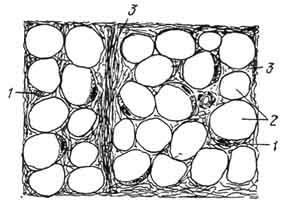
Структурная единица белой жировой ткани - шаровидные крупные (до 120 мкм в диаметре) зрелые жировые клетки с характерным микроскопическим строением (рис. 109). Большая часть объема клетки занята одной крупной каплей жира. Овальное ядро и цитоплазма находятся на периферии клетки. Такая клетка при световой микроскопии гистологического среза, окрашенного с применением жирорастворяющих веществ, имеет
156
Рис. 109. Схема строения клеток белой жировой ткани:
1 - ядро жировой клетки; 2 - полость, оставшаяся после растворения капли жира; 3 - соединительная ткань.
перстеневидную форму. В результате растворения жира на месте жировой капли в клетке остается светлая вакуоль. При электронной микроскопии в околоядерной зоне обнаруживают главным образом удлиненные митохондрии, другие органеллы слабо выражены. При развитии клеток жировые включения в цитоплазме появляются сначала в виде мелких рассеянных капель, позднее сливающихся в одну крупную каплю. Жировые вещества в клетках можно выявить при использовании специальных красителей (судан III, судан IV, четырехокись осмия).
Из жировых клеток в жировой ткани образованы дольки различных размеров и формы. Между дольками находятся прослойки рыхлой соединительной ткани, в которых проходят мелкие кровеносные сосуды и нервные волокна. Между жировыми клетками внутри долек
располагаются отдельные клетки соединительной ткани (фиброциты, тканевые базофилы), сеть тонких аргирофильных волокон и кровеносные капилляры.
Общее количество белой жировой ткани в организме животных различных видов, пород, пола, возраста, упитанности колеблется от 1 до 30% к живой массе. Запасные жиры
вжировой ткани - это наиболее высококалорийные вещества, при окислении которых в организме освобождается большое количество энергии (1 г жира =39 кДж). Подкожная жировая ткань, особенно у диких животных, имеет большое значение для защиты организма от механических повреждений, предохраняет от потерь тепла. Жировая ткань вдоль сосудисто-нервных пучков, в капсуле и оболочках органов обеспечивает их относительную изоляцию, защиту и ограничение подвижности. Скопления жировых клеток
всочетании с окружающими их пучками коллагеновых волокон в коже подошв и лап создают хорошие амортизационные свойства. Значительна роль жировой ткани как депо воды. Образование воды - важная особенность обмена жиров у животных, обитающих в засушливых районах (верблюды).
При голодании организм мобилизует прежде всего запасные жиры из клеток жировых депо. В них уменьшаются и исчезают жировые включения.
Жировая ткань глазной орбиты, эпикарда, лап сохраняется даже при сильном истощении.
Цвет жировой ткани зависит от вида, породы и типа кормления животных. У большинства животных, за исключением
157
свиней и коз, в жире содержится пигмент каротин, придающий желтый цвет жировой ткани.
Б у р а я ж и р о в а я т к а н ь в значительном количестве имеется у грызунов и животных, впадающих в зимнюю спячку, а также у новорожденных животных других видов. Расположена преимущественно под кожей между лопатками, в шейной области, в средостении и вдоль аорты.
Состоит она из относительно мелких клеток, очень плотно прилегающих друг к другу, напоминая внешне железистую ткань. К клеткам подходят многочисленные волокна симпатической нервной системы, они оплетены густой сетью кровеносных капилляров. Для клеток бурой жировой ткани в сравнении с клетками белой жировой ткани характерны центрально расположенное ядро и наличие в цитоплазме мелких жировых капель, слияния которых в более крупную каплю не происходит. В промежутках между жировыми каплями расположены многочисленные митохондрии и значительное количество гранул гликогена. Содержащиеся в митохондриях окрашенные белки системы транспорта электронов - цитохромы придают бурый цвет этой ткани.
В клетках бурой жировой ткани интенсивно идут окислительные процессы, сопровождаемые выделением значительного количества энергии. Однако большая часть образующейся энергии расходуется не на синтез молекул АТФ, а на теплообразование. Такое свойство липоцитов бурой ткани является важным для регуляции температуры у новорожденных животных и согревания животных после пробуждения от зимней спячки.
Пигментные клетки (пигментоциты), как правило, отростчатой формы. В
цитоплазме много темно-коричневых или черных зерен пигмента из группы меланинов.
Значительное количество пигментных клеток - хроматофоров в соединительной ткани кожи низших позвоночных - рептилий, амфибий, рыб, у которых они обусловливают ту или иную окраску внешнего покрова и выполняют защитную функцию. У млекопитающих пигментные клетки сосредоточены преимущественно в соединительной ткани стенки глазного яблока - склере, сосудистой и радужной оболочках, а также в ресничном теле.
М е ж к л е т о ч н о е в е щ е с т в о рыхлой соединительной ткани составляет значительную ее часть. Представлено оно коллагеновыми и эластическими волокнами, расположенными относительно рыхло и беспорядочно, и основным (аморфным) веществом. В межклеточном веществе осуществляются разнообразные ферментативные обменные процессы, перемещение различных веществ и клеточных элементов, самосборка и перестройка волокон в соответствии с направлением действия механических факторов. В межклеточном веществе расположены чувствительные нервные окончания, непрерывно посылающие в центральную нервную систему сигналы о его состоянии.
Коллагеновые волокна - основные волокна, обеспечивающие механическую прочность ткани. В рыхлой соединительной ткани
158
они имеют вид лентовидных тяжей, ориентированных в различных направлениях. Волокна не ветвятся, им свойственна малая растяжимость, большая прочность на разрыв (выдерживают до 6 кг на 1 мм2 поперечного сечения), способность объединяться в пучки. При длительной варке коллагеновые волокна образуют клей (колла), откуда и произошло название волокон.
Прочность коллагеновых волокон обусловлена их более тонкой структурной организацией. Каждое волокно состоит из фибрилл диаметром до 100 нм, расположенных параллельно друг другу и погруженных в межфибриллярное вещество, содержащее гликопротеиды, гликозаминогликаны и протеогликаны. Под электронным микроскопом по длине фибриллы наблюдают характерную поперечную полосчатость - чередование темных
исветлых полос с определенным периодом повторяемости, а именно олив темный и один светлый сегмент вместе составляют один период длиной 64 - 70 нм. Наиболее отчетливо эта полосчатость видна на негативно окрашенных препаратах коллагеновых фибрилл. При электронной микроскопии позитивно окрашенных фибрилл, кроме основной темносветлой периодичности, выявляют сложный рисунок более тонких электроноплотных полосок, разделенных узкими промежутками шириной 3 - 4 hm.
Внастоящее время характерный рисунок строения коллагеновой фибриллы объясняют специфичностью ее макромолекулярной организации. Фибрилла состоит из более тонких микрофибрилл, образованных молекулами белка тропоколлагена. Последние имеют длину 280 - 300 нм и ширину 1,5 нм и являются своеобразными мономерами (рис. 110). Образование фибриллы - результат характерной группировки мономеров в продольном и поперечном направлении. Мономеры уложены параллельными рядами и удерживаются друг около друга ковалентными поперечными связями, причем в одном ряду между концами соседних мономеров имеется зазор, равный 0,4 длины периода, а по ширине мономеры одного ряда накладываются на мономеры соседнего со смещением на 1/4 его длины. Такое чередование зазоров и наложений создает полосчатый вид фибрилл на электронных микрофотограммах. Одна молекула тропоколлагена пересекает пять светлых
ичетыре темных сегментов (рис. 111).

Известно также, что по длине молекула тропоколлагена асимметрична и там, где сходные последовательности аминокислот оказываются друг против друга, возникают узкие вторичные темноокрашенные полосы. Каждая молекула тропоколлагена представляет собой спираль из трех полипептидных цепей, удерживаемых друг около друга водородными связями. Уникальная структура тропоколлагена обусловлена особенно высоким содержанием в нем глицина (до 30%), а также оксилизина и оксипролина. В зависимости от аминокислотного состава и формы объединения цепей в тройную спираль различают четыре основных типа коллагена, имеющих различную локализацию в организме. Коллагены I типа является наиболее распространенным и содержится в
159
Рис. 110. Схема строения коллагенового волокна:
А - спиральная структура макромолекулы коллагена (по Ричу); мелкие светлые кружки - глицин; крупные светлые кружки - пролин; заштрихованные кружки - гидроксипролин; Б - схема строения коллагеновых волокон; 1 - пучок фибрилл; 2 - фибрилла; 3 - протофибрилла; 4 - молекула коллагена.
Рис. 111. Коллагеновая фибрилла:
А - электронная микрофотография негативно окрашенной коллагеновой фибриллы (ув. 180000); Б - схема расположения тропоколлагеновых молекул, объясняющая возникновение поперечной исчерченности (по Ходжа и Петруски, 1964): 1 - темные сегменты соответствуют промежуткам между концами молекул тропоколлагена; 2 - светлые сегменты соответствуют зонам перекрывания молекул.
соединительной ткани кожи, сухожилий и в костях. Коллаген 11 типа имеется преимущественно в гиалиновом и волокнистом хрящах. В коже зародышей, стенке кровеносных сосудов, связках преобладает коллаген III типа, а в базальных мембранах -