

цитология гистология и эмбриология александровская козлов радостина
.pdf9 :: 10 :: 11 :: 12 :: Содержание
КРАТКИЙ ИСТОРИЧЕСКИЙ ОЧЕРК
Первые микроскопические исследования строения тканей растений, как и термин "клетка", принадлежат физику Роберту Гуку (1665). Позднее клеточное строение растительных, а затем и животных тканей описывали M. Мальпиги (1671 - 1675), H. Грю (1671), А. Левенгук (1673 - 1695), Сваммердам (1737) и др. Левенгук, работая с микроскопами своей конструкции (лупами), дающими увеличение до 300 раз, описал эритроциты крови, их движение по капиллярам, спермии, строение поперечнополосатых мышечных волокон, нервные волокна, простейшие микроорганизмы и многое другое.
Несмотря на успешность микроскопических исследований в XVII и в начале XVIII вв., они не привлекли к себе особого внимания и не получили широкого развития. Это определялось как низким качеством микроскопов, страдающих сферической и хроматической аберрациями, так и влиянием господствующей в то время теории преформации (Галлер), утверждающей, что ничто в природе не возникает заново и развитие организмов - это
9
только процесс развертывания зачатков, заложенных при сотворении мира. При развитии новых организмов происходит лишь рост сформированного и заложенного в половую клетку (яйцевую клетку или спермий) организма.
Как реакция па теорию преформации в XVIII в. пришла теория эпигенеза (Вольф, 1749 - 1769), утверждающая, что организм всегда развивается из бесструктурного вещества яйцевой клетки путем новообразования органов. К. Ф. Вольф в 1760 г. по приглашению Петербургской Академии наук переехал из Германии в Россию, где плодотворно работал до конца своей научной деятельности. Он первым наблюдал образование органов из "листовидных пластинок" (зародышевых листков). Кроме того, Вольф изучил и описал развитие сердца у цыпленка, почек и др.
Первым в России применившим в научных исследованиях микроскоп был M. В. Ломоносов. По его инициативе при Петербургской Академии наук созданы оптические мастерские, сыгравшие значительную роль в успешном развитии в России естественных наук. Основным недостатком микроскопов того времени была хроматическая аберрация, препятствующая четкому выявлению структур.
Крупным успехом микроскопии, определившим ее дальнейшее развитие, было создание ахроматического микроскопа. Теоретическая разработка последнего выполнена петербургским академиком Л. Эйлером, а его учеником, академиком H. Фуссом, в 1877 г. сделаны вычисления конструктирования ахроматических линз. Работу по созданию ахроматического микроскопа завершил академик Ф. У. Эпинус, сконструировавший его первую модель (1784). Позднее он дал новую, более совершенную модель такого микроскопа, но она, к сожалению, была изготовлена только в двух экземплярах. Один из них в настоящее время находится в коллекции Академии Наук СССР. Одновременно ахроматические микроскопы разрабатывались и в Голландии.
С 30-х годов XVIII в. началось фабричное производство микроскопов. В России в XVLII в. микроскопические исследования, начало которым положил M. В. Ломоносов, проводили И. Кулеман, изучавший яичник овец в процессе полового цикла и беременности, Петр Аш,
анализировавший сперму, К. Ф. Вольф, описавший развитие кишечника кур, и A. M. Шумлянский - микроскопическое строение почек. Микроскоп успешно использовался и в учебном процессе. Введение в практику научных исследований ахроматических микроскопов обеспечило успешность изучения растительных и животных тканей, а соответственно и внимание к результатам исследований.
В числе пионеров современного микроскопического анализа следует прежде всего назвать Я. Пуркине (Бреславль) и его учеников.
Я. Пуркине, успешно разрабатывая технику микроскопических исследований и анализируя клетки ("комочки") тканей различных органов животных, впервые описал ядро в яйцевых клетках курицы ("зародышевый пузырек") и в нервных клетках (1825 - 1827). Несколько позднее описание ядра было сделано и в растительных клетках (Браун, 1831). Микроскопические исследования привлекли внимание многих ученых, таких как Г. Валентин, А. Дютраше, Л. Ф. Горянинов, Я. Генле, P. Ремак, M. Шлейден, T. Шванп и многие другие. В числе исследователей, наиболее близко стоящих к формулировке клеточной теории, был Пуркине, но он и его ученики (Валентин и др.), описывая в различных тканях животных клетки, а в ряде случаев и их ядра, не анализировали своих материалов в свете общности структурной организации животных и растений.
Честь создания клеточной теории принадлежит T. Шванну (1838 - 1839), который показал, что клетки тканей животных и растительных организмов принципиально сходны, они гомологичны друг с другом по развитию и строению и аналогичны по функциональному значению. T. Шванн, анализируя свои наблюдения образования клеток животных и сопоставляя их с аналогичным процессом растительных тканей (Шлейден), пришел к выводу, что в основе строения как животных, так и растительных тканей лежат клетки, а ядра являются признаком их развития. Это дало основание Шванну оцепить клеточное строение как всеобщую закономерность, характеризующую единство органической природы - животных и растительных организмов.
10
Ф. Энгельс определил клеточную теорию как величайшее открытие века, сопоставив ее значение с двумя другими великими открытиями: законом сохранения энергии и эволюционной теорией Дарвина.
Важную роль в развитии клеточной теории Шванна имели труды патолога P. Вирхова (1858), что отражено в его афоризме OMNIS CELLULA ET CELULLA (всякая клетка происходит только от клетки). Вирхов в своих исследованиях показал, что в основе патологических процессов (воспаления, дистрофии, патологических новообразований и др.) лежат изменения клеток.
Клеточная теория, сформулированная Шванном под влиянием Вирхова, распространилась на патологию и медицину и была принята как основная теория, объясняющая нормальные и патологические процессы живой природы.
В России гистологические исследования в XIX в. принимают систематический характер. Организуются самостоятельные кафедры гистологии на медицинских факультетах университетов в Москве и Петербурге (1868), а позднее в Казани, Киеве, Харькове, Дерпте (Тарту), что определило формирование соответствующих гистологических коллективов (школ), имеющих определенные научные направления. Так, исследования микроскопического строения центральной и периферической нервной
системы H. M. Якубовичем позволило дифференцировать различные виды клеток коры головного мозга. Кафедру гистологии Московского университета возглавил А. И. Бабухин (1827 - 1891). Под его руководством успешно разрабатывались вопросы развития и функции органов нервной системы, сетчатки глаза, электрического органа рыб и др. И. Ф. Огнев (1855 - 1928) изучал влияние на организм различных внешних и внутренних факторов.
В Петербурге в университете и медико-хирургической академии кафедры гистологиивозглавляли Ф. В. Овсянников (1827 - 1906), H. M. Якубович (1817 - 1879), А. С. Догель
(1852 - 1922), M. Д. Лавдовский (1846 - 1903), А. А. Максимов (1874 - 1928), А. А. Заварзин
(1886 - 1945), И. Г. Хлопин (1897 - 1961) и др. Ими и их учениками и последователями (Немилов, Данинц, Хлокин, Румянцев, Ясвоин, Елисеев, Кадилов и др.) собран и обобщен огромный материал сравнительно-гистологических и экспериментальных исследований соединительной и эпителиальной тканей. Разрабатывались вопросы закономерностей эмбрионального гистогенеза (Кацнельсон, Щелкунов, Винников, Кноре), структурной организации эндокринной системы, процессов гистогенеза и регенерации мышечной ткани (Немилов, Румянцев, Алешин, Студитский и др.).
Организатором кафедры гистологии в Киевском университете был П. П. Неремежко (1868). Исследования гистологов этой кафедры были направлены па анализ эмбрионального развития зародышевых листков и органов зародыша: глаз, надпочечников, селезенки, печени, щитовидной и поджелудочной желез, кровеносных сосудов, мышечной ткани и др.
Нейрогистологические исследования активно проводились в Казани К. А. Арнштейном, А. С. Догелем, A. E. Смирновым, Д. А. Тимофеевым, а позднее A. H. Миславским и Б. И. Лаврентьевым и их учениками H. Г. Колосовым, И. Ф. Ивановым, Г. И. Забусовым, E. К. Плечковой, M. А. Григорьевой, П. А. Ковальским и многими другими.
Вопросы структурной и гистохимической организации тканей и органов сельскохозяйственных животных в настоящее время успешно изучают коллективы гистологов под руководством Ю. T. Техвера, О. В. Александровской, Л. В. Давлетовой, П. А. Ильина, А. Ф. Рыжих, И. G. Ржаницыной, H. А. Гороховского, А. И. Пилипенко, Л. П. Тельцова и др. Особое внимание исследователей привлекают гистохимический, биохимический и электронно-микроскопический анализы тканей и органов животных организмов.
Клеточная теория в ее современном виде включает следующие положения: 1) клетка - это наименьшая единица живого. Данное представление было сформулировано T. Шванном и развито в трудах P. Вирхова. Существование симпластов и синцитиев (см. ниже) не опровергает этого положения; 2) клетки различных тканей различных организмов гомологичны по своему строению, то есть, несмотря на их большое многообразие и специфические особенности, все клетки всех организмов имеют общий принцип строения:
11
они имеют ядро, цитоплазму, основные органеллы. Это положение клеточной теории было также сформулировано T. Шванном; 3) размножение клеток происходит только путем деления исходной клетки; 4) клетки рассматривают как части целостного организма, они специализированы, имеют определенные функции и структуру, взаимосвязаны в функциональных системах тканей, органов, систем органов.

К числу так называемых неклеточных структур относятся симпласты и синцитии. Их существование, естественно, не опровергает положения клеточной теории о клетке как структурной единице живого, так как возникают эти структуры либо от слияния клеток, либо в результате деления ядер без последующей цитотомии. С и м п л а с т ы - это многоядерные структуры, состоящие из большого объема цитоплазмы, включающей множество ядер. Примером симпласта может служить поперечнополосатое мышечное волокно. С и н ц и т и и (соклетия) - клетки, связанные цитоплазматическими перемычками. В настоящее время установлено, что большинство структур, которые до развития электронной микроскопии считались синцитиями (например, ретикулярная ткань органов кроветворения), в действительности являются клетками, соприкасающимися своими отростками. Однако некоторые клетки (сперматогонии) при делении сохраняют связь цитоплазматическими перемычками, образуя группы в несколько клеток.
12
9 :: 10 :: 11 :: 12 :: Содержание

12 :: 13 :: 14 :: Содержание
ГЛАВА 1
ОСНОВЫ ОБЩЕЙ ЦИТОЛОГИИ
Цитология - наука о развитии, строении и жизнедеятельности клеток. Клетки являются основным структурным и функциональным элементом организма. Их форма, размеры и специфичность дифференцировки разнообразны, характерны для различных тканей и в значительной мере отражают своеобразие их организации в связи со специфичностью их функций. Так, клетки крови, взвешенные в ее плазме, округлые. Клетки, выстилающие поверхность, плотно прилежат друг к другу и имеют плоскую, кубическую или призматическую форму. Клетки гладкой мышечной ткани вытянутые, веретенообразные. У нервных клеток длинные отростки, что позволяет им проводить свои импульсы на большие расстояния (рис. 4).
Вещество клетки - протоплазма - в процессе жизнедеятельности непрерывно взаимодействует с окружающей средой. Химический состав ее определяется специфичностью обмена веществ организма. Известно, что 96% массы животного составляют 4 элемента: углерод, кислород, водород и азот. В значительных количествах (в сумме до 3%) в тканях содержатся калий, кальций, натрий, фосфор, сера, магний, железо, хлор. Все остальные химические элементы, входящие в состав тканей организма, - микроэлементы (медь, марганец, кобальт, цинк и др.) - содержатся в сотых и тысячных долях процента, участвуют в важных физиологических процессах, имеют существенное значение в жизнедеятельности организма.
Химические элементы входят в состав протоплазмы в виде сложных органических соединений - белков, углеводов, липидов, нуклеиновых кислот и др.
12
Рис. 4. Форма и общий принцип строения фиксированных клеток (схема):
1 - цилиндрические клетки эпителия кишечника; 2 - кубические клетки мочевых канальцев почки; 3 - плоские клетки мезотелия брюшины; 4 - округлые клетки крови (а - с дольчатым ядром - нейтрофильный лейкоцит, б - с округлым ядром - лимфоциты); 5 - веретеновидная клетка с палочковидным ядром (гладкая мышечная клетка); 6 - отростчатая (нервная) клетка; 7 - бокаловидная клетка эпителия кишечника; 8 - клетка с ресничками (из многорядного мерцательного эпителия дыхательных путей); 9 - крылатая (сухожильная) клетка; 10 - жгутиковая клетка (спермий); и - многоядерная клетка (остеокласт); 12 - безъядерные клетки (эритроциты).
По определению В. Я. Александрова, клетка представляет собой живую систему, состоящую из двух важнейших, неразрывно связанных между собой частей - цитоплазмы и ядра.
Наиболее распространенный компонент структурной организации клетки - биологические мембраны. В их составе локализованы различные субстанции и энзимы, катализирующие многочисленные специфические, характерные для клеток химические реакции, протекающие на границе двух фаз: между структурами цитоплазмы - органеллами и цитоплазматическим матриксом, между клетками и окружающей средой. Мембраны регламентируют взаимодействие ферментов и субстратов во времени.
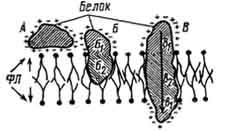
При значительном разнообразии строения клеточных мембран все они представлены пластами липопротеидной природы (липиды - 40%, белки - 60%). Молекулы липидов биологических мембран характеризуются наличием несущих заряд полярных головок - гидрофильного полюса молекулы и неполярных хвостов (их гидрофобного полюса), образованных жирными кислотами. Взаимодействие последних формирует жидкостнобимолекулярный слой липидов биологических мембран. Белки в составе мембраны связываются с липидами как с помощью ионных, так и на основе гидрофобных связей, погружаясь в липидный слой мембраны (рис. 5).
13
Рис. 5. Взаимодействие белков с липидными слоями:
А - белковая молекула, связанная ионными взаимодействиями; Б, В - гидрофильные (б1, в1) и гидрофобные (б2, B2) взаимодействия белков с липидами (фл) (по Покровскому и Тутельману).
Мембранные белки представлены тремя разновидностями: периферическими, интегральными и полу интегральными.
Периферические белки располагаются на поверхности мембраны. Их молекулы связаны с полярными головками молекул лппи-дов электростатическим взаимодействием. Интегральные и полуинтегральные белки погружены в липидный слой. Молекулы интегральных белков проходят весь липидный слой мембраны. Их гидрофобная часть находится в средине молекулы и соответственно локализуется в гидрофобной зоне липидной фазы мембраны. В молекулах полуинтегральных белков гидрофобные аминокислоты сосредоточены на одном полюсе, в соответствии с этим они погружаются в липидный слои мембраны лишь наполовину - полюсом, взаимодействующим с гидрофобной частью липидных молекул.

Белки мембран, взаимодействуя с молекулами липидов, не закреплены жестко и способны менять степень погружения в липидный слой и перемещаться в плоскости мембраны.
На мембранах фиксируются ансамбли различных ферментов. Они участвуют в реакциях, протекающих на границе между органеллами и цитоплазматическим матриксом и между клеткой и окружающей ее средой. Мембраны регулируют время реакции, скорость активного транспорта субстратов и энзимов, обеспечивают высокую степень химической неоднородности, характерную для цитоплазмы клеток.
14
12 :: 13 :: 14 :: Содержание
14 :: 15 :: 16 :: 17 :: 18 :: 19 :: Содержание
ЯДРО
Ядро - обязательная составная часть полноценной клетки. Оно содержит геном и продуцирует макромолекулы, контролирующие синтетические процессы цитоплазмы. Клетки без ядра (эритроциты млекопитающих, кровяные пластинки, центральные волокна хрусталика) не способны продуцировать белок и соответственно ограничены в метаболической активности. Форма ядер и их размеры в клетках различного типа весьма разнообразны и специфичны. Для большинства плоских, кубических и округлых клеток характерна шарообразная форма ядра. Такую же форму имеют отростчатые нервные клетки, тогда как в числе шарообразных клеток крови имеются клетки с сегментированными ядрами (рис. 6).
В ядре различают ядерную оболочку, хроматин, ядрышко и ядерный сок.
14
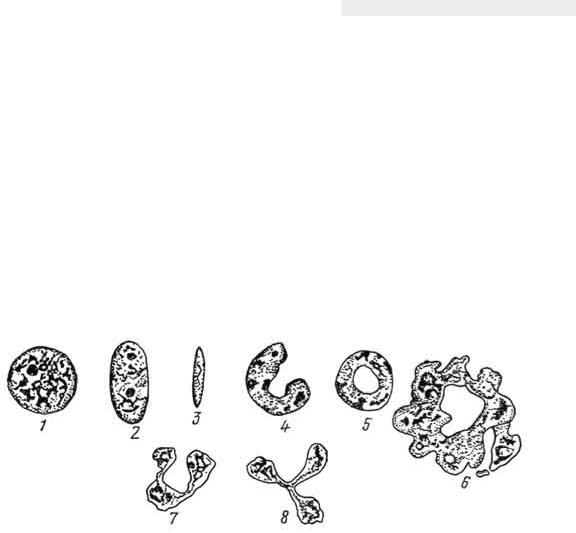
Рис. 6. Форма ядер в различных клетках:
1 - круглая, пузырьковидная; 2 - овальная; 3 - палочковидная; 4 - подковообразная; 5 - кольцевидная; 6 - неправильно-кольцевидная; 7 - подковообразная; S - трехлопастная.
Ядерная оболочка (кариолемма) состоит из двух элементарных липопротеидных мембран, разделенных п е р и н у к л е а р н ы м пространством в 20 - 100 нм (рис. 7 и 8). На определенном расстоянии мембраны ядерной оболочки формируют ядерные поры диаметром 80 - 90 нм. В области поры наружная и внутренняя мембраны ядерной оболочки сближаются и непосредственно переходят одна в другую. Количество пор зависит от функциональной активности клетки. В мембраны пор включены три ряда гранул по восемь гранул в каждом, расположенные последовательно со стороны ядра, в центре поры и со стороны цитоплазмы клетки. Каждая гранула снабжена фибриллой. Последние при определенном положении, перпендикулярном к стенке поры, в совокупности образуют "д и а ф р а г м у ", закрывающую пору (рис. 9). Электронно-микроскопическим анализом установлен непосредственный переход наружной мембраны ядерной оболочки в мембраны эндо-плазматической сети цитоплазмы, что, очевидно, необходимо учитывать при оценке возможных путей переноса веществ через ядерную оболочку.
Хроматин ядра получил свое название за способность интенсивно окрашиваться основными красителями (CHROMA - краска), что зависит главным образом от присутствия в ядре дезоксирибонуклеиновой кислоты (ДНК). Последняя является характерной составной частью хромосом, определяющей закономерности основных жизненных процессов в клетках. Химический анализ хромосом свидетельствует, что выделенное из их ядер
вещество (дезоксирибонуклеопротеид - ДНП) содержит ДНК (40% общей массы), незначительное количество РНК (до 1%) и белки (60%, 85% гистонов и 15% кислых белков).
Молекула ДНК представляет собой двойную спираль нуклеотпдов. Специфичность структуры молекул ДНК и РНК, характерная для каждого организма, определяется содержанием и последовательностью локализации в их составе нуклеотидов, различных по азотистым основаниям. Химический состав и строение ДНК,
15
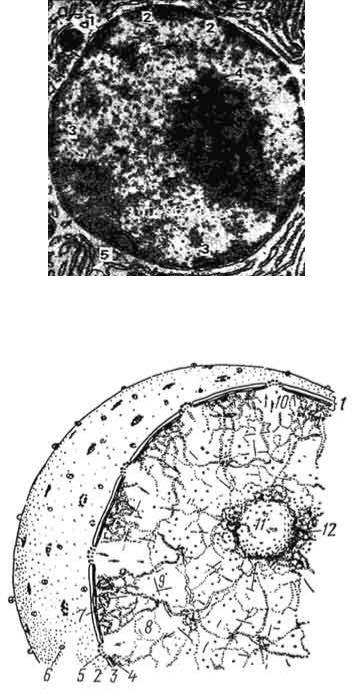
Рис. 7. Электронная микрофотография ядра кроветворной клетки (ув. 16000):
1 - оболочка ядра; 2 - поры в оболочке; 3 - глыбки хроматина; 4 - ядрышко; 5 - гранулярная сеть (по Фаусету).
Рис. 8. Схема интерфазного ядра клетки (по Заварзину и Хазаровой):
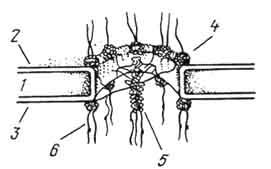
1 - поверхностный аппарат ядра; 2, 3 - мембраны ядерной оболочки (2 - наружная, 3 - внутренняя, между ними перинуклеарное пространство); 4 - плотная пластика; 5 - поровой комплекс; 6 - рибосомы; 7 - гетерохроматин; 8 - эухроматин (ДЫП); 9 - элементы ядерного матрикса; 10 - РШТ-частицы; 11 - ядрышко; 12 - околоядерный хроматин.
PиC. 9. Тонкая организация ядерной поры:
1 - перинуклеарное пространство; 2 - внутренняя ядерная мембрана; 3 - внешняя ядерная мембрана; 4 - периферические субъединицы; 5 - центральная гранула (по Франке).
16
механизм ее синтеза и синтеза РНК подробно излагаются в курсе генетики.
Из хроматина построены хромосомы. Однако хромосомы как палочковидные или нитевидные структуры видны только в определенные фазы деления клеток. В ядрах неделящихся клеток значительные участки каждой хромосомы деконденсированы или деспирализованы. Эти д и с п е р с н ы е , или растянутые, участки хромосом активны в процессах синтеза. Они плохо окрашиваются гистологическими красителями. Дисперсный хроматин называют э у х р о м а т и н о м . Участки хромосом, которые остаются конденсированными, интенсивно окрашиваются и выглядят на гистологических препаратах как базофильные глыбки в ядре. Они функционально не активны.
Конденсированный хроматин в ядре неделящейся клетки называют г е т е р о х р о м а т и н о м .
Белки хромосом представлены гистонами и негистоновыми белками.
Гистоны характеризуются высоким сродством к ДНК и образуют с ней характерные структурные комплексы. В составе хромосом различают пять фракций гистонов: H1, Н2а, Н2б, Н3, Н4. Четыре из них (Н2а, Н2б, Н3 и Н4 по две молекулы каждого вида) формируют глобули - н у к л е о с о м ы - 20 нм в диаметре. Участки молекул ДНК в два витка спирали (140 нуклеотидных пар молекулы) стабилизируются нуклеосомами. Нить ДНК, расположенная между нуклеосомами, имеет различную длину и состоит из 10 - 70 пар оснований. Молекула гистона HL связана с этими сегментами хромосомы и участвует в установлении и стабилизации высокоупорядоченной структуры хромосомы (рис. 10).
Растянутая молекула ДНК нуклеосомы и промежуточных участков составляют филаменты (10 нм в диаметре). В присутствии ионов магния филаменты конденсируются с участием гистона H1 в структурные комплексы высшего порядка диаметром 20 - 30 нм и более.
Микроскопическая картина хроматина ядра клеток соответствует степени напряженности синтетических процессов определенных участков хромосом. При активизации последних хромосомы деспирализуются и рассредоточиваются - базофильная