

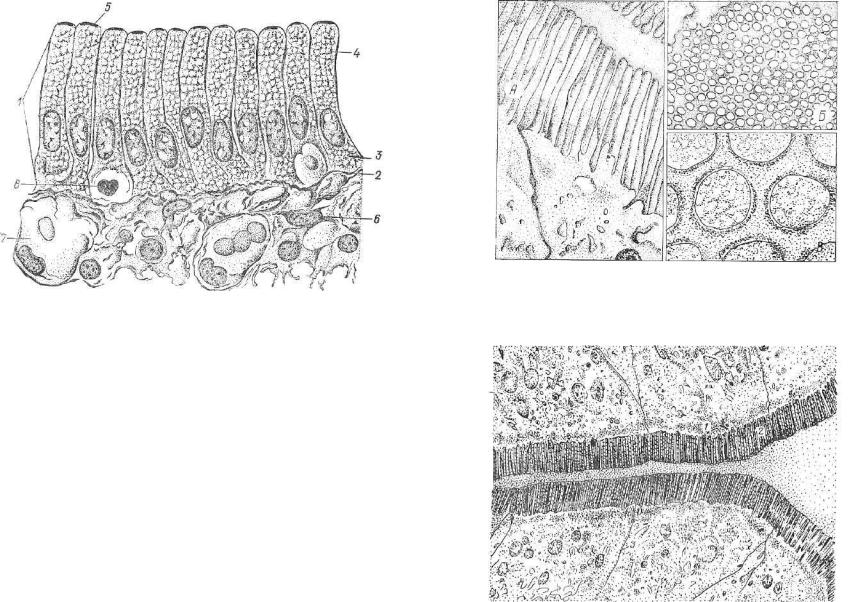
gistologia
.pdfб — однослойный кубический; |
в — однослойный цилиндрический |
(столбча |
|||||
тый); |
г — однослойный многорядный цилиндрический мерцательный (псев |
||||||
домногослойный); г—1 — мерцательная клетка; г—2 — мерцательные реснич |
|||||||
ки; г—з — вставочные (замещающие) клетки; |
д — многослойный |
плоский |
|||||
(сквамозный) неороговевающий; |
д—i — клетки базального |
слоя; |
#—2 — |
||||
клетки шиповатого слоя; д—8 — клетки поверхностного слоя; |
е — многослой |
||||||
ный плоский (сквамозный) ороговевающий эпителий; е—а — базальный слой; |
|||||||
е—б — шиповатый слой; е—в — зернистый слой; е—г — блестящий слой; е— |
|||||||
д — роговой слой; ж — переходный эпителий; |
ж—а — клетки |
базального |
|||||
слоя; |
ж—б — клетки промежуточного |
слоя; |
ж—в — клетки |
покровного |
|||
слоя; |
з — рыхлая соединительная ткань; |
и — бокаловидная клетка. |
|
||||
пания, пальцевидных соединений. Овальные ядра эпителиоцитов обычно сдвинуты к базальному полюсу и расположены на одной высоте от базальной мембраны.
Модификации простого столбчатого эпителия — каемчатый эпителий кишечника (рис. 81) и железистый эпителий желудка (см. гл, 11). Покрывая внутреннюю поверхность слизистой обо лочки кишечника, каемчатый эпителий участвует во всасывании питательных веществ. Все клетки этого эпителия, называемые микроворсинчатыми эпителиоцитами, расположены на базальной мембране. В этом эпителии хорошо выражена полярная дифференция, которая обусловлена строением и функцией его эпителиоцитов. Полюс клетки, обращенный в просвет кишечника (апикаль ный полюс), покрыт исчерченной каемкой. Под ней в цитоплазме расположена центросома. Ядро эпителиоцита лежит в базальном полюсе. Комплекс Гольджи прилегает к ядру, рибосомы, мито хондрии и лизосомы рассредоточены по всей цитоплазме.
Таким образом, в апикальном и базальном полюсах микровор синчатого эпителиоцита находятся разные внутриклеточные струк туры, это и называется п о л я р н о й д и ф ф е р е н ц и а ц и е й .
Клетки кишечного эпителия называются микроворсинчатыми, так как на их апикальном полюсе расположена исчерченная каем ка — слой микроворсинок, образованный выростами плазмолеммы апикальной поверхности эпителиоцита. Микроворсинки отчетливо
105
1 — эпителиальная клетка; 2 — базальная мембрана; 3 — базальный полюс; 4 — апикальный полюс; 5 — исчерченная каемка; б"^— рыхлая соединительная ткань; 7 — кровеносный сосуд; 8 — лейкоцит.
различимы только в электронный микроскоп (рис/82, 83). Каждый эпителиоцит имеет в среднем более тысячи микроворсинок. Они увеличивают всасывающую поверхность клетки, а следовательно,
икишечника до 30 раз.
Вэпителиальном пласте этого эпителия находятся бокаловид ные клетки (рис. 84). Это одноклеточные железы, вырабатываю щие слизь, которая предохраняет клетки от вредных воздействий механических и химических факторов.
Простой столбчатый железистый эпителий покрывает внутрен нюю поверхность слизистой оболочки желудка. Все клетки эпите лиального пласта расположены на базальной мембране, их высо та больше ширины. В клетках четко представлена полярная диф ференциация: овальное ядро и органеллы находятся на базальном полюсе, в апикальном — лежат капли секрета, отсутствуют органеллы (см. гл. 10).
Однослойный, однорядный цилиндрический мерцательный эпителий (псевдомиогослойный реснитчатый эпителий) (рис. 85) выстилает воздухоносные пути органов дыхания — носовую по лость, гортань, трахею, бронхи, а также канальца придатка семен ника, внутреннюю поверхность слизистой оболочки яйцевода. Эпителий воздухоносных путей развивается из энтодермы, эпите лий органов воспроизводства — из мезодермы.
Рис. 82. А — микроворсинки исчерченной каемки и примыкающий к ней участок цитоплазмы эпителиоцита (ув. 21800, продольное сечение); Б — поперечное сечение микроворсинок (ув. 21800); В — поперечное сечение микроворсинок (ув. 150 000). Электронная микрофотография.
1 — апикальный полюс эпителиоцита; 2 — всасывающая каемка; з —* плазмолемма эпителиоцита. Электронная микрофотография.
(106

Рис. 84. Бокаловидные клетки:
1 — клетки эпителия; 2 — бокаловид* ные клетки в начальной стадии образо» вания секрета; з — бокаловидные клет* ни, выделяющие секрет; 4 — ядро; Д— секрет.
Все клетки эпителиального пласта лежат на базальной мем бране, отличаются по форме, строению, функции. В эпителии] воздухоносных путей располага ются и бокаловидные клетки; свободной поверхности достига ют только реснитчатые цилинд рические и бокаловидные клет ки. Между ними вклиниваются стволовые (замещающие) эпителиоциты. Высота и ширина этих
клеток варьируют: некоторые из них столбчатой формы, их овальные ядра находятся в центре клетки; другие более низкие с расши ренным базальным и суженным апикальным полюсами. Округлые ядра расположены ближе к базальной мембране. Все разновидно сти вставочных эпителиоцитов не имеют мерцательных ресничек.
Следовательно, ядра цилиндрических реснитчатых, замещаю щих и низких замещающих клеток расположеньКрядами на раз ной высоте от базальной мембраны, в связи с чем эпителий назы вают многорядным. Псевдомногослойным (ложномногослойным)1 его именуют потому, что все эпителиоциты находятся на базаль ной мембране.
Между мерцательными и вставочными (замещающими) клет ками лежат одноклеточные железы — бокаловидные клетки, про дуцирующие слизь. Она накапливается в апикальном полюсе, от тесняя к основанию клетки эндоплазматическую сеть, комплекс Гольджи, митохондрии и ядро. Последнее при этом приобретает форму полулуния, очень богато хроматином и интенсивно окра шивается. Секрет бокаловидных клеток покрывает эпителиаль ный пласт и способствует прилипанию вредных частиц, микроор ганизмов, вирусов, попавших в воздухоносные пути вместе с вды хаемым воздухом.
Мерцательные (реснитчатые) эпителиоциты — высоко диффе ренцированные клетки, поэтому митотически неактивны. На своей поверхности у реснитчатой клетки около трехсот ресничек, каждая из которых образована тонким выростом цитоплазмы, покрытым плазмолеммой. В ресничке содержится одна центральная пара и девять пар периферических микротрубочек. У основания реснич* ки периферические микротрубочки исчезают, а центральная про* ходит вглубь, образуя базальное тельце.
Базальные тельца всех эпителиоцитов расположены на одном уровне (рис. 86). Реснички находятся в постоянном движении. Их направление движения будет перпендикулярно плоскости за легания центральной пары микротрубочек. Благодаря движению ресничек из органов дыхания удаляются попавшие с воздухом пы линки и избыточное скопление слизи. В половых органах мерцание ресничек способствует продвижению яйцеклеток.
109
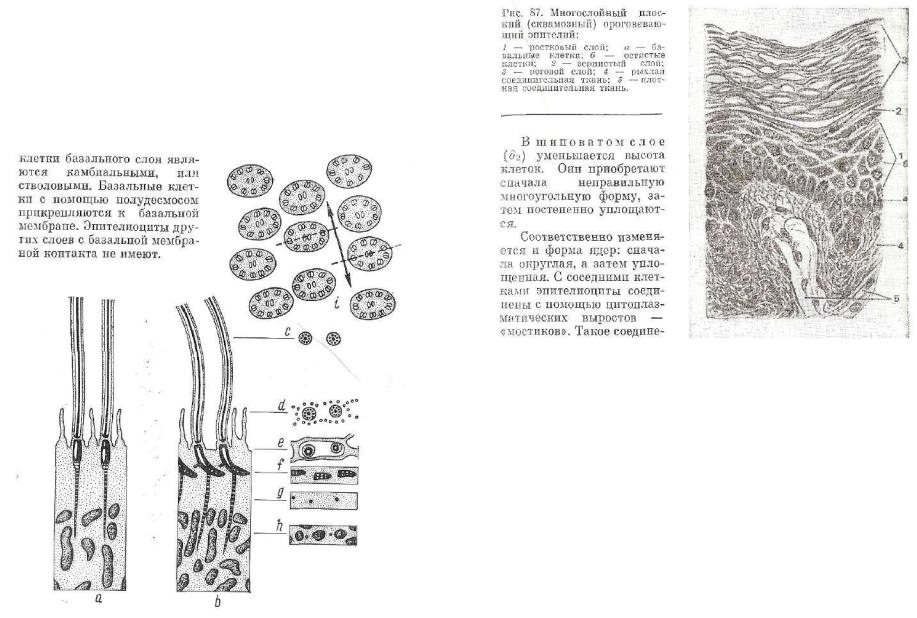
Рис. 86. Схема реснитчатого аппарата эпителия:
а — разрез в плоскости, перпендикулярной к плоскости движения реснички; Ь — раэрез в плоскости движения реснички; с—Л — поперечное сечение ресничек на разных уровнях; г — поперечный разрез ресничек {пунктиром показана плоскость, перпендикулярная к направлению движения).
ПО
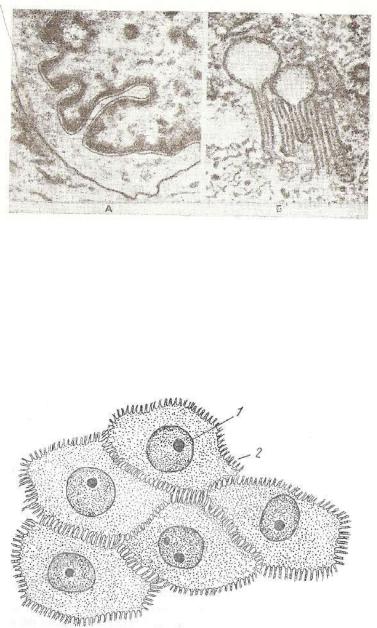
плоским эпителием. В нем также различают базальный, шипова тый, плоский слои клеток. /
Все клетки базального слоя (см. рис. 79, е—а) расположены на базальной мембране. Большинство клеток этого слоя называ ются кератиноцитами. Имеются и другие клетки — меланйциты и беспигментные гранулярные дендроциты (клетки Лангерганса). Кератиноциты участвуют в синтезе волокнистых белков, полиса харидов, липидов. Они имеют столбчатую форму, их ядра бога ты ДНК, а цитоплазма — РНК. В клетках содержатся также тон кие нити — тонофибриллы, зерна пигмента меланина.
Кератиноциты базального слоя обладают максимальной митотической активностью. После митоза часть дочерних клеток пе ремещается в располоя?енный выше шиповатый слой, другие — в виде «запаса» остаются в базальном слое, выполняя функцию камбиальных (стволовых) эпителиоцитов. Основное значение кератиноцитов — образование плотного, защитного, неживого, рогового вещества — кератина, что обусловило название клеток.
Меланиноциты отросчатой формы. Их клеточные тела распо ложены в базальном слое, а отростки могут достигать других сло ев эпителиального пласта. Основная функция меланоцитов — образование меланосом и кожного пигмента меланина. Послед ний по отросткам меланоцита может передаваться другим клеткам эпителия. Кожный пигмент предохраняет организм от чрезмерно го ультрафиолетового облучения, негативно влияющего на орга низм. Ядра меланоцитов занимают большую часть клетки, непра вильной формы, богаты хроматином. Цитоплазма светлее, чем у кератиноцитов, в ней много рибосом, развиты гранулярная эндоплазматическая сеть, аппарат Гольджи. Эти органеллы принима ют участие в синтезе меланосом, которые овальной формы и со стоят из нескольких плотных гранул, покрытых мембраной.
Безпигментные |
(светлые) гранулярные |
дендроциты имеют |
2—5 отростков. В |
их цитоплазме содержатся |
особые гранулы, |
сходные по форме с теннисной ракеткой (рис. 88). Значение этих клеток не выяснено. Существует мнение, что их функция связана с контролем пролиферативной активности кератиноцитов.
Клетки шиповатого слоя не связаны с базальной мембраной. Они многогранной формы; перемещаясь к поверхности, постепен но уплощаются. Граница между клетками обычно неровная, так как на поверхности кератиноцитов формируются цитоплазматические выросты («шипики»), при помощи которых они соединяют ся друг с другом. Это приводит к образованию клеточных мости ков (рис. 89) и межклеточных щелей. По межклеточным щелям протекает тканевая жидкость, содержащая питательные вещества и ненужные продукты обмена веществ, предназначенные для уда ления. В клетках этого слоя очень хорошо развиты тонофибрил лы. Их диаметр равен 7—10 нм. Располагаясь пучками, они за канчиваются в зонах десмосом, прочно соединяющих клетки друг с другом при формировании эпителиального пласта. Тонофибрил лы выполняют функцию опорно-защитного каркаса.
Рис. 88. А — клетка Лангерганса; В — специфические гранулы «теннисные ракетки с ампулярным концевым расширением и продольными ламеллами в области рукоятки». Электронная микрофотография.
З е р н и с т ы й с л о й (см. рис. 79, е—в) состоит из 2—4 ря дов клеток плоской формы, лежащих параллельно поверхности эпителиального пласта. Для эпителиоцитов характерны округлые, овальные или вытянутые ядра; уменьшение количества органелл; накопление кератиногиалинового вещества, пропитывающего то нофибриллы. Кератогиалин окрашивается основными красителя ми, поэтому имеет вид базофильных гранул. Кератиноциты
Рис. 89. Клеточные мостики в эпидермисе носового зеркальца быка:
112

/
зернистого слоя являются предшественниками клеток следующе го — б л е с т я щ е г о с л о я (е—г). Его клетки лишены ядер и органелл, а тонофибриллярно-кератиногиалиновые комплексы сливаются в гомогенную массу, сильно преломляющую свет и окрашивающуюся кислыми красителями. Электронно-микроскопи- чески этот слой не выявлен, так как не имеет ультраструктурных отличий.
Р о г о в о й слой (е—д) состоит из роговых чешуек. Они об разуются из блестящего слоя и построены из кератиновых фиб рилл и аморфного электроноплотиого материала, роговой слой снаружи покрыт однослойной мембраной. В поверхностных зонах фибриллы лежат более плотно. Роговые чешуйки соединяются друг с другом с помощью ороговевших десмосом и других струк тур клеточных контактов. Потеря роговых чешуек возмещается новообразованием клеток базального слоя.
Итак, кератиноциты поверхностного слоя превращаются в плот* ное неживое вещество — кератин (keratos — рог). Он защищает нижележащие живые клетки от сильных механических воздейст вий и высыхания. Кератин препятствует вытеканию тканевой жидкости из межклеточных щелей.
Роговой слой выполняет функцию первичного защитного барь ера, так как он непроницаем для микроорганизмов. Ороговевающий плоский и многослойный эпителий может достигать значи тельной толщины, что приводит к нарушению питания его клеток. •Это устраняется образованием соединительнотканных вырос тов — сосочков, которые увеличивают поверхность контакта кле ток базального слоя и рыхлой соединительной ткани, выполняю щей трофическую функцию.
Переходный эпителий {ж) развивается из мезодермы и по крывает внутреннюю поверхность почечной лоханки, мочеточников, мочевого пузыря. При функционировании этих органов меняется объем их полостей, в связи с чем толщина эпителиального пласта то резко снижается, то возрастает.
Эпителиальный пласт состоит из базального, промежуточного, поверхностного слоев {ж—а, б, в).
Базальный слой построен из базальных клеток, связанных с базальной мембраной, различных по форме и размеру: мелкие кубические и крупные грушевидные клетки. Первые из них имеют округлые ядра и базофильную цитоплазму. В эпителиальном плас те ядра этих клеток образуют самый нижний ряд ядер. Мелкие кубические клетки характеризуются высокой митотической актив ностью и выполняют функцию стволовых клеток. Вторые — своей узкой частью прикреплены к базальной мембране. Их расширен ное тело расположено над кубическими клетками; цитоплазма светлая, так как слабо выражена базофилия. Если орган не на полнен мочой, крупные грушевидные клетки нагромождаются друг на друга, формируя как бы промежуточный слой.
Покровные клетки уплощены. Часто многоядерны или их ядра полиплоидны (содержат большее количество хромосом по сравне-
Ш
Рис. 90. Переходный эпителий почечной лоханки овцы:
а — а' — слизистая клетка покровной зоны со слабой реакцией на сливь; б — промежуточная зона; в — митоз; г — базальная зона; д •— соединительная ткань.
чевом пузыре.
нию с диплоидным набором хромосом). Поверхностные клерки могут ослизняться. Эта способность особенно хорошо развита у травоядных (рис. 90). Слизь предохраняет эпителиоциты от вред ных воздействий мочи.
Таким образом в перестройке эпителиального пласта данного зида эпителия играет степень наполнения органа мочой (рис. 91).
ЖЕЛЕЗИСТЫЙ ЭПИТЕЛИЙ
Способность клеток организма интенсивно синтезировать актив ные вещества (секрет, гормон), необходимые для осуществления функции других органов, характерна для эпителиальной ткани. Эпителии, вырабатывающие секреты, называются железистыми, а его клетки — секреторными клетками, или секреторными гландулоцитами. Из секреторных клеток построены железы, которые могут быть оформлены в виде самостоятельного органа или яв ляться только его частью.
Различают эндокринные (endo — внутри, krio — отделяю) и экзокринные (ехо — снаружи) железы. Э к з о к р и н н ы е желе зы состоят из двух частей: концевой (секретирующей) части и выводных протоков, по которым секрет поступает на поверхность организма или в полость внутреннего органа.. Выводные протоки •обычно не принимают участие в образовании секрета.
Эндокринные железы лишены выводных протоков. Их ак тивные вещества (гормоны) поступают в кровь, в связи с чем функцию выводных протоков выполняют капилляры, с которыми железистые клетки очень тесно связаны. Подробно функциональ ная морфология желез внутренней секреции будет рассмотрена в главе 8.
Экзокринные железы разнообразны по строению и функции. Они могут быть одноклеточными и многоклеточными. Примером одноклеточных желез служат бокаловидные клетки, встречающие ся в простом столбчатом каемчатом и псевдомногослойном реснит чатом эпителиях. Несекретирующая бокаловидная клетка цилин дрической формы и сходна с несекреторными эпителиоцитами. Секрет (муцин) накапливается в апикальной зоне, а ядро и органеллы смещаются к базальной части клетки. Смещенное ядро при обретает форму полулуния, а клетка — бокала. Затем секрет из ливается из клетки, а она вновь приобретает столбчатую форму.
Экзокринные многоклеточные железы могут быть однослой ными и многослойными, что обусловлено генетически. Если желе за развивается из многослойного эпителия (потовая, сальная. молочная, слюнные железы), то и железа многослойна; если из однослойного (железы дна желудка, матки, поджелудочная желе за), то они однослойны.
Характер ветвления выводных протоков экзокринных желез
.различен, поэтому они подразделяются на простые и сложные. У простых желез неветвящийся выводной проток, у сложных — ветвящийся.
Концевые отделы у простых желез разветвляются и не развет вляются, у сложных — разветвляются. В связи с этим у них и соответствующие названия: разветвленная железа и неразветвлен-
ная железа.
По форме концевых отделов экзокринные железы классифици руют на альвеолярные, трубчатые, трубчато-альвеолярные. У аль веолярной железы клетки концевых отделов формируют пузырь ки или мешочки, у трубчатых — образуют вид трубочки. Форма концевой части трубчато-альвеолярной железы занимает проме жуточное положение между мешочком и трубочкой (рис. 92, 93).
117
116
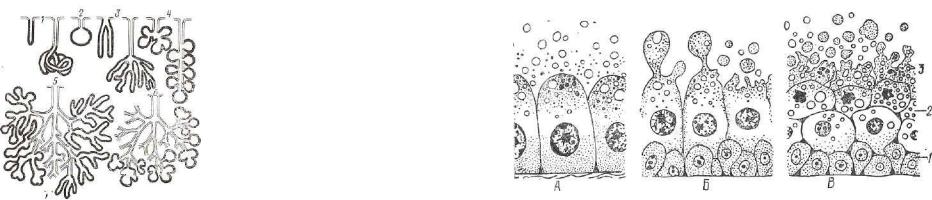
Рис. 93. Схематическое изображениепростых и сложных экзокринных /же лез:
1 — простые трубчатые железы с нераз- |
|||||||
ветвленными концевыми отделами; г — |
|||||||
простая альвеолярная железа с нераз- |
|||||||
ветвленным концевым |
отделом; |
з — |
|||||
простые трубчатые железы с разветвлен |
|||||||
ными концевыми |
отделами; |
4 |
— |
про |
|||
стые |
альвеолярные |
железы |
с |
раавет- |
|||
вленными |
концевыми |
отделами; |
S — |
||||
сложная |
альвеолярно-трубчатая |
железа |
|||||
с разветвленным концевым отделом; б— |
|||||||
сложная альвеолярная железа с развет |
|||||||
вленными |
концевыми |
отделами. |
Секре |
||||
торные |
отделы изображены черными ли |
||||||
ниями, |
выводные |
ьротокн — |
светлыми. |
||||
Клетки концевого отдела именуются г л а н д у л о ц и т а м и . Процесс синтеза секрета начинается с момента поглощения гландулоцитами из крови и лимфы исходных компонентов секрета. При активном участии органелл, синтезирующих секрет белково го или углеводного характера, в гландулоцитах образуются секре торные гранулы. Они накапливаются в апикальной части клетки,, а затем путем обратного пиноцитоза выделяются в полость кон цевого отдела. Завершающий этап секреторного цикла — восста новление клеточных структур, если в процессе секреции они раз рушились.
Строение клеток концевой части экзокринных желез обуслов лено составом выделяемого секрета и способом его образования.
По способу образования секрета железы делят на голокринные, апокринные, мерокринные (зккринные). При г о л о к р и н - н о й секреции (holos — целый) железистый метаморфоз гландулоцитов начинается с периферии концевого отдела и протекает в направлении выводного протока. Примером голокринной секре ции является сальная железа. Стволовые клетки с базофильной цитоплазмой и округлым ядром расположены на периферии кон цевой части. Они интенсивно делятся митозом, поэтому мелкие по размеру. Перемещаясь к центру железы, секреторные клетки уве личиваются, так как в их цитоплазме постепенно накапливаются капельки кожного жира. Чем больше откладывается в цитоплазме жировых капель, тем интенсивнее протекает процесс деструкции органелл. Он завершается полным разрушением клетки. Плазмолемма разрывается, а содержимое гландулоцита поступает в про свет выводного протока.
При а п о к р и н н о й секреции (аро — от, сверху) разрушает ся апикальная часть секреторной клетки, являясь затем состав ной частью ее секрета. Данный тип секреции совершается в пото вой или молочной железах.
При м е р о к р и н н о й секреции клетка не разрушается. Такой способ образования секрета типичен для многих желез организма: железы желудка, слюнные железы, поджелудочная железа, эндо кринные железы (рис. 94).
А — мерокринный; Е — апокринный; В — голокринный; 1 — маяодиФФеренпированные клетки; 2 — перерождающиеся клетки; 3 — разрушающиеся клетки.
Таким образом, железистый эпителий так же, как и покров ный, развивается из всех трех зародышевых листков (эктодермы, мезодермы, энтодермы), расположен на соединительной ткани, лишен кровеносных сосудов, поэтому питание осуществляется диффузионным способом. Клеткам свойственна полярная дифференцировка: в апикальном полюсе локализуется секрет, в база ль ном полюсе — ядро и органеллы.
Регенерация. Покровные эпителии занимают пограничное по ложение. Они часто повреждаются, поэтому характеризуются вы сокой регенерационной способностью. Регенерация осуществляет ся главным образом митомическим и очень редко амитотическим способом. Клетки эпителиального пласта быстро изнашиваются, стареют и гибнут. Их восстановление называется ф и з и о л о г и ч е с к о й р е г е н е р а ц и е й .
Восстановление эпителиальных клеток, утраченных по причи не травмы и дрзтой патологии, называется р е п а р а т и в н о й
ре г е н е р а ц и е й .
Воднослойных эпителиях регенерационной способностью об ладают или все клетки эпителиального пласта, или, если эпителиоциты высокодифференцированны, то за счет зонально лежа щих своих стволовых клеток.
Вмногослойных эпителиях стволовые клетки находятся на базальной мембране, поэтому лежат в глубине эпителиального пласта.
Вжелезистом эпителии характер регенерации обусловлен спо собом образования секрета. При голокринной секреции стволовые клетки находятся снаружи железы на базальной мембране. Делясь
идифференцируя.сь, стволовые клетки преобразуются в желези стые.
Вмерокринных и апокринных железах восстановление эпителиоцитов протекает главным образом путем внутриклеточной ре генерации.
118
ТКАНИ ВНУТРЕННЕЙ СРЕДЫ (ОПОРНО-ТРОФИЧЕСКИЕ ТКАНИ)
Ткани внутренней среды возникают одновременно с эпителиаль ными на самых ранних этапах развития многоклеточных живот ных. У высших позвоночных они представлены группой тканей,, общим морфологическим признаком которых является наличие в их составе не только клеток, но и хорошо развитого межклеточно го вещества. В соответствии со специфичностью и различием кле точного состава и в большей степени особенностями структурной организации межклеточного вещества среди тканей внутренней среды выделяют: кровь и лимфу, разновидности соединительных тканей, хрящевую и костную ткани. Проявлением единства этих видов тканей при резком различии физико-химических свойств (кровь и лимфа —жидкие, костная ткань — твердая) является происхождение их из общего эмбрионального источника — мезен химы.
Всем тканям внутренней среды свойственны трофическая и защитная, а тканям соединительным, хрящевым и костным — в той или иной степени механическая и опорная функции.
МЕЗЕНХИМА
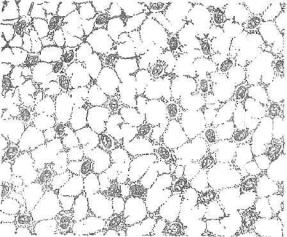
Мезенхима — совокупность эмбриональных сетевидно связанных: отростчатых клеток, заполняющих промежутки между более ком пактными эпителиоподобными зародышевыми листками и зачат ками органов. В ячеях этой сети находится студенистое межкле точное вещество (рис. 95).
Рис. 95. Мезенхима.
При эмбриогенезе раньше всего мезенхима появляется в со ставе внезародышевых органов. Подтверждается это тем фактом, что первые кровяные островки возникают в стенке желточного мешка. В теле зародыша мезенхима возникает главным образом из клеток определенных участков мезодермы — дерматомов, склеротомов и спланхнотомов. В области головы часть мезенхимы раз вивается из клеток, выселяющихся из эктодермальной ганглиозной пластинки, — нейромезенхима. Клетки мезенхимы быстро делятся митозом. В различных ее участках возникают многочис ленные мезенхимные производные — кровяные островки с их эн дотелием и клетками крови, клетки соединительных тканей и гладкой мышечной ткани, образуются уплотненные клеточные за чатки скелетных тканей и др.
«КРОВЬ
Внутрисосудистая кровь — подвижная тканевая система с жид ким межклеточным веществом — плазмой и форменными элемен тами — эритроцитами, лейкоцитами и кровяными пластинками (тромбоцитами — у птиц и низших позвоночных).
Гистогенетически, структурно и функционально сосудистая кровь является частью системы крови и тесно связана с органами кроветворения и кроверазрушения, рыхлой соединительной тканью и другими тканями и органами. Многие лейкоциты цир кулируют в крови непродолжительное время (несколько дней), находятся в ней в относительно недеятельном состоянии и явля ются предшественниками клеток, активная специфическая дея тельность которых осуществляется после выхода этих лейкоцитов из кровотока в составе тканей (преимущественно рыхлой соедини тельной ткани) и органов.
Эритроциты и кровяные пластинки выполняют свои функции непосредственно в кровяном русле. В капиллярном отделе сосу дистой системы происходят интенсивный обмен между составны ми частями плазмы крови и окружающей тканевой жидкости и миграция форменных элементов крови.
Постоянно циркулируя в замкнутой системе кровообращения, кровь объединяет работу всех систем организма и поддерживает многие физиологические показатели внутренней среды организма на определенном, оптимальном для осуществления обменных про цессов уровне. На основе циркуляции форменных элементов и со ставных веществ плазмы кровь выполняет в оргапизме разносто ронние жизненно важные функции: дыхательную, трофическую, защитную, регуляторную, выделительную и другие. Конкретное понимание многочисленных функций крови возможно лишь на основе изучения строения и свойств ее основных компонентов — форменных элементов и, плазмы.
Несмотря на подвижность и изменяемость крови, ее показа тели в каждый момент соответствуют функциональному состоя-
121
120
нию организма, поэтому исследование крови является одним из важнейших диагностических методов.
Плазма — жидкая составная часть крови, содержит 90—92% воды и 8—10% сухих веществ, в составе которых около 9% орга нических и 1% минеральных веществ. Основные органические вещества плазмы крови — белки (альбумины, различные фракции глобулинов и фибриноген). С белками крови связано онкотическое давление, имеющее существенное значение в процессах транс капиллярного обмена между составными частями плазмы крови и тканевой жидкостью. Иммунные белки (антитела), а большин ство их содержится в 7-гл°булиновой фракции, называют имму
н о г л о б у л и н а м и (Ig). Альбумины |
обеспечивают перенос |
различных веществ — свободных жирных |
кислот, билирубина и |
др. Фибриноген принимает участие в процессах свертывания кро ви. Более полные сведения о составе и свойствах плазмы крови приводятся в курсах биохимии и физиологии.
Форменные элементы крови
Эритроциты
Эритроциты (erythros — красный) — высокоспециализированные клетки, приспособленные для выполнения основной функции кро ви — транспорта кислорода и углекислого газа в организме. В 1 мкл крови у позвоночных содержится несколько миллионов
эритроцитов, а у большинства |
сельскохозяйственных |
животных |
|||
от 5 до 10 млн (табл. 1). |
|
|
|
|
|
1. Количество эритроцитов в крови животных |
|
|
|||
|
|
|
|
|
|
Виды животных, |
Эритроциты, |
|
Виды ЖИВОТНЫХ, |
Эритроциты, |
|
в том числе и птиц |
млн/мкл |
|
в том числе И птиц |
млн/мкл |
|
Крупный рогатый скот |
5,0— 7,5 |
|
Собаки |
5,2— |
8,4 |
Лошади |
6,0— 9,0 |
|
Кошки |
6,6— 9,4 |
|
Овцы |
7,0—12,0 |
|
Кролики |
4,5— 7,5 |
|
Козы |
12,0—18,0 |
|
Куры |
3,0— 4,0 |
|
Северные олбни |
6,5— 8,5 |
|
Утки |
3,0- |
4,5 |
Свиньи |
6,0- 7,5 |
|
Индейки |
2,5- |
3,5 |
Определение количества эритроцитов в крови — важная со ставная часть общего клинического анализа крови животных, оно проводится либо с помощью счетной камеры, либо в электронных автоматических счетчиках. Количество эритроцитов в крови зави сит от вида, породы, возраста животных и может изменяться под влиянием различных факторов — физической нагрузки, баромет рического давления, а также при болезнях.
Утратив в процессе развития ядро, зрелые эритроциты у мле копитающих являются безъядерными клетками и имеют форму двояковогнутого круглого диска со средним диаметром круга
5—7 мкм. Эритроциты крови верблюда и ламы овальной формы. Дисковидная форма увеличивает общую поверхность эритроцита в 1,64 раза по сравнению с поверхностью шара такого же диамет ра, что способствует ускорению проникновения кислорода в эрит роцит. Эритроциты других позвоночных — птиц, рептилий, амфи бий и рыб — овальной формы, имеют ядро с сильно конденсиро ванным хроматином. Они крупнее эритроцитов млекопитающих (например, у саламандр величина их превышает в 100 раз).
Б большинстве случаев между количеством эритроцитов и их величиной можно обнаружить обратную зависимость: например, у ноз в 1 мкл крови 14 млн эритроцитов, диаметр эритроцита 4 мкм; у лягушки в 1 мкл крови 0,35 млн эритроцитов, диаметр овального эритроцита по длине 22,8 мкм, а по ширине 15,8 мкм. У живот ных одного вида все эритроциты почти одинаковой величины и появление в крови эритроцитов другой величины и формы счита ется признаком патологического процесса.
Эритроциты покрыты оболочкой — плазмолеммой (толщиной около 6 нм), содержащей 44% липидов, 47% белков и 7% угле водов. Многие мембранные белки эритроцитов являются гликопротеидами и гликолипидами, их поверхностные концевые олигосахаридные компоненты определяют групповые свойства крови. Мембрана эритроцитов легко проницаема для газов, анионов, обеспечивает активный перенос ионов натрия, облегченный транс порт глюкозы. Внутреннее коллоидное содержимое эритроцитов на 34% состоит из г е м о г л о б и н а — уникального сложного окрашенного соединения — хромопротеида, в небелковой части которого (геме) имеется двухвалентное железо, способное обра зовывать особые непрочные связи с молекулой кислорода. Имен но благодаря гемоглобину осуществляется дыхательная функция эритроцитов. При высокой концентрации кислорода, особенно в капиллярах легких, происходит присоединение молекул кислоро да к атомам железа — образуется о к с и г е м о г л о б и н .
При низкой концентрации кислорода в капиллярах других ор ганов связи между кислородом и железом легко разрываются и кислород отсоединяется — образуется восстановленный гемогло бин, придающий венозной крови синевато-вишневый цвет. Таким образом, функционирование эритроцитов осуществляется непо средственно в сосудистой крови. Обладая большой суммарной по верхностью, эритроциты, кроме транспортировки газов, участвуют в переносе адсорбированных на их оболочке различных веществаминокислот, ферментов и др.
Наличие гемоглобина в эритроцитах обусловливает выражен ную оксифилию их при окраске мазка крови по Романовскому— Гимзе (смесь кислого красителя эозина и основного — азура II). Эритроциты при этом окрашиваются в красный цвет эозином. Так как эритроциты имеют форму двояковогнутого диска, то централь ная часть клетки окрашивается слабее, чем периферическая. Нор мальными по окраске считаются эритроциты, центральная часть которых составляет около трети диаметра эритроцита. При неко-
122 |
123 |